§ 3. Строение клетки
Размеры клетки широко варьируют от 0,1 мкм (некоторые бактерии) до 155 мм (яйцо страуса). У всех клеток, независимо от их формы, размеров, функциональной нагрузки обнаруживается сходное строение (рис. 3.13).
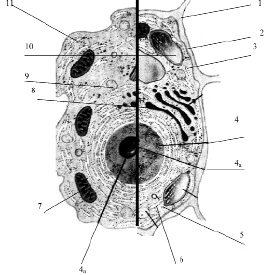
Рис. 3.13. Схема строения живой клетки: 1 – оболочка; 2 – мембрана; 3 – цитоплазма; 4 – ядро; 4а – ядрышко; 5 – рибосомы; 6 – эндоплазматическая сеть (ЭПС); 7 – митохондрии; 8 – комплекс гольджи; 9 – лизосомы; 10 – пластиды; 11 – клеточные включения
Снаружи клетка одета мембраной. Внутренняя часть клетки содержит многочисленные органоиды – структурные образования клетки, выполняющие определенные функции жизнедеятельности клетки.
1. Оболочка. Присутствует только у растительных клеток. Состоит из волокон целлюлозы. Функции оболочки: защита клетки от внешних повреждений, придает стабильную форму клетки, эластичность растительным тканям.
Повреждение наружной оболочки приводит к гибели клетки (цитолиз).
2. Мембрана. Тончайшая структура (75 Ǻ), состоит из двойного слоя молекул липидов и одного слоя белков. Такая структура обеспечивает уникальную эластичность и прочность мембране

участие в обмене веществ. Эта функция связана с избирательной проницаемостью в клетку определенных веществ и выведение из нее продуктов обмена. В процессе питания в клетку могут проникать определенные растворы веществ (пиноцитоз) и твердые частицы (фагоцитоз).
Явление фагоцитоза – поглощение клеткой твердых частиц – впервые было описано русским врачом Мечниковым. Фагоцитарная особенность лежит в основе процесса иммунитета. Особенно развита у лейкоцитов, клеток костного мозга, лимфатических узлов, селезенки, надпочечников и гипофиза.
Пиноцитоз – поглощение клеткой растворов – состоит в том, что мельчайшие пузырьки жидкости втягиваются через образующуюся воронку, проникают через мембрану и усваиваются клеткой.
3. Цитоплазма – внутренняя среда клетки. Представляет собой гелеобразную жидкость (коллоидная система), состоит на 80 % из воды, в которой растворены белки, липиды, углеводы, неорганические вещества. Цитоплазма живой клетки находится в постоянном движении (циклоз).




все биохимические реакции происходят только в водных растворах, что обеспечивается в среде цитоплазмы.
4. Ядро – обязательный органоид эукариотических клеток. Впервые было исследовано и описано Р. Броуном в 1831 г. В молодых клетках расположено в центре клетки, в старых – смещается в сторону. Снаружи ядро окружено мембраной с крупными порами, способными пропускать крупные макромолекулы. Внутри ядро заполнено клеточным соком – кариоплазмой, основная часть ядра заполнена хроматином – ядерным веществом, содержащим ДНК и белок. Перед делением хроматин образует палочковидные хромосомы. Причём, хромосомы одинакового строения (но содержащие разные ДНК!) образуют пары, зрительно воспринимаемые как одно целое (рис. 3.14).

Рис. 3.14. Хромосомный набор человеческой клетки перед началом деления
Структурирование всех хромосом в пары свидетельствует о том, что число хромосом – чётное. Поэтому, его часто обозначают 2n, где n – количество хромосомных пар, а соответствующий набор хромосом называют диплоидным. Например, у голубей n = 40 (80 хромосом), у мухи n = 6 (12 хромосом), у собаки n = 39 (78 хромосом), у аскариды n = 1 (2 хромосомы). У человека n = 23 (46 хромосом). Однако, в половых клетках число хромосом в два раза меньше. Поэтому набор хромосом в половых клетках называется гаплоидным. Клетки, не являющиеся половыми называются соматическими. Иногда клетки с гаплоидным набором хромосом называют гаплоидными клетками, а с диплоидным набором хромосом – диплоидными клетками.
При слиянии двух родительских гаплоидных половых клеток образуется диплоидная клетка, дающая начало новому организму с набором генов отца и матери
Совокупность всех хромосом ядра (а значит и генов) клетки называется генотип. Именно генотип определяет все внешние и внутренние признаки конкретного организма.
В соматических клетках 44 Х-образные хромосомы (22 пары) у женщин и мужчин идентичны (сходны по строению), их называют аутосомами. А 23-я пара имеет конфигурацию ХХ – у женщин и ХY – у мужчин. Эти пары хромосом именуются половыми хромосомами.
Необходимо помнить, что хотя в соматических клетках набор хромосом диплоидный (2n), однако, перед началом деления клеток происходит репликация ДНК, то есть, удвоение их количества, а, значит, и удвоение количества хромосом. Поэтому перед началом деления соматической клетки в ней насчитывается 4n хромосом (рис. 16). Она становится тетраплоидной.
– хранение генетической информации;
– контроль за всеми процессами, происходящими в клетке: делением, дыханием, питанием и др.
4а. Ядрышко – структура, содержащаяся в ядре. Ядро может содержат 1, 2 или более ядрышек. Функция ядрышка – формирование рибосом.
Следует отметить, что не все клетки имеют оформленное ядро. Клетки, имеющие ядро называются эукариотическими или эукариотами. Клетки, не имеющие ядра, называются прокариотическими или прокариотами. Функции ядра у прокариот несёт одна нить ДНК (именуется хромосома), в которой хранится вся генетическая информация. К прокариотам относятся бактерии и сине-зеленые водоросли. Как правило, у прокариотов отсутствуют и некоторые другие органоиды. Размеры прокариотических клеток меньше, чем размеры эукариот.
5. Рибосомы – самые мелкие органоиды клетки. Были обнаружены в 1954 г. Французским ученым Паладом. Рибосомы были обнаружены в цитоплазме, а также на гранулярной ЭПС и в ядре.
Функция рибосом: обеспечение биосинтеза белка.
6. Эндоплазматическая сеть. Представляет собой каналы и полости, ограниченные мембраной. Различают две разновидности ЭПС: гранулярная ЭПС и агранулярная ЭПС. Гранулярная ЭПС морфологически отличается от агранулярной наличием на ее поверхности многочисленных рибосом (на агранулярной ЭПС рибосомы отсутствуют).
Функции эндоплазматической сети:
– участие в синтезе органических веществ: на гранулярной ЭПС синтезируются белки, на агранулярной – липиды и углеводы;
– транспортировка продуктов синтеза ко всем частям клетки.
Несложно уяснить, что гранулярная ЭПС характерна для клеток, синтезирующих белки (например клетки желез внутренней секреции), агранулярная ЭПС характерна для клеток-производителей углеводов и липидов (например клетки жировой ткани).
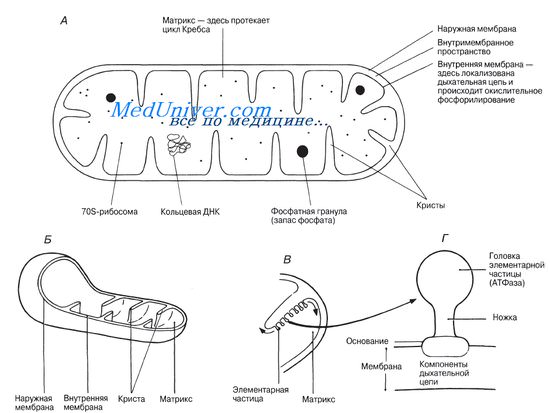
7. Митохондрии – крупные органоиды, состоящие из двойного слоя мембран: наружная – гладкая, внутренняя образует многочисленные гребнеобразные складки – кристы. Внутри митохондрии заполнены жидкостью (матрикс).
Функции митохондрий: основная функция митохондрий – обеспечение клетки энергией. Этот процесс происходит за счет синтеза аденозинтрифосфорной кислоты (АТФ) (рис. 3.15), в которой фрагмент

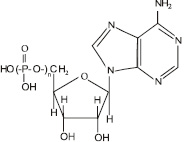
Рис. 3.15. Структурная формула аденозинфосфорных кислот. Для аденозинтрифосфорной кислоты n = 3, для аденозиндифосфорной кислоты n = 2, для аденозинмонофосфорной кислоты n = 1
При взаимодействии молекулы аденозинтрифосфорной кислоты с водой отщепляется один остаток фосфорной кислоты, в результате чего образуется аденозиндифосфорная кислота – АДФ и выделяется огромное количество энергии:
Впоследствии от АДФ может отщепляться еще один остаток фосфорной кислоты, образуя АМФ – аденозинмонофосфорную кислоту.
Освободившаяся энергия используется для жизнедеятельности клетки (КПД процесса превышает 80 %!).
Наряду с распадом АТФ и выделением энергии в клетке постоянно происходит синтез АТФ и накопление энергии (обратные реакции).
Количество митохондрий в клетке зависит от потребности последней в энергии. Так, в клетках кожи человека находится в среднем 5–6 митохондрий, в клетках мышц – до 1000, в клетках печени – до 2500!
8. Комплекс Гольджи. Итальянский ученый Гольджи обнаружил и описал структуру клетки, напоминающую стопки мембран, цистерны, пузырьки и трубочки. Расположена эта система чаще всего возле ядра.
Функции комплекса Гольджи: в полостях комплекса накапливаются всевозможные продукты обмена клетки, которые по каким-либо причинам не вывелись наружу. В последствии эти продукты могут быть использованы клеткой для процессов жизнедеятельности. Из пузырьков и цистерночек комплекса Гольджи в растительных клетках образуются вакуоли, заполненные клеточным соком.
9. Лизосомы – мелкие органоиды. Представляют собой пузырьки, окруженные мембраной. Внутри лизосомы заполнены пищеварительными ферментами (обнаружено 12 ферментов), которые расщепляют и переваривают крупные макромолекулы (белки, полисахариды, нуклеиновые кислоты).
Функции лизосом: растворение и переваривание макромолекул. Лизосомы участвуют в фагоцитозе. Понятно, что основная функция по перевариванию поступающих в клетку частиц принадлежит лизосомам.
10. Пластиды. Эти органоиды характерны только для растительных клеток. Форма напоминает двояковыпуклую линзу. Структура пластид напоминает таковую у митохондрий: двойной слой мембраны. Наружная – гладкая, внутренняя образует складки, называемые тилакоидами. На тилакоидах происходит основной жизненно важный для всех зеленых растений процесс – фотосинтез:

Пластиды бывают трех типов:
1) Хлоропласты – зеленые пластиды. Их цвет обусловлен наличием хлорофилла. Хлорофилл – основное вещество хлоропластов (имеет зеленый цвет). Только благодаря хлорофиллу возможен процесс фотосинтеза (см. раздел 4.2). Хлоропласты придают зеленый цвет растительным организмам.
2) Хромопласты – пластиды, имеющие различные окраски: от ярко-желтого до пурпурно-багряного. Наличие различных пигментов окрашивают плоды, цветки и осенние листья растений в соответствующие цвета. Этот факт особенно важен для привлечения насекомых к цветкам, как природный индикатор созревания плодов и др.
3) Лейкопласты – бесцветные пластиды, в которых происходит накопление запасных питательных веществ (например, крахмала).
Некоторые виды пластид могут переходить друг в друга: например, переход хлоропластов в хромопласты: созревание томатов, яблок, вишни, и т. д.; изменение окраски листьев в осенний период времени. Лейкопласты могут переходить в хлоропласты: позеленение картофеля на свету. Это доказывает общность происхождения пластид.
11. Клеточные включения. Вакуоли. Это непостоянные и необязательные составляющие клетки. Они могут появляться и исчезать в течение всей жизни клетки. К ним относятся капли жира, зерна крахмала и гликогена, кристаллы щавелево-кислого кальция и др. Жидкие продукты обмена называются клеточным соком и накапливаются они в вакуолях. В клеточном соке растворены сахара, минеральные соли, пигменты и т. д. Чем старше клетка, тем больше клеточного сока накапливает клетка. Молодые клетки практически не содержат вакуолей.
Помимо перечисленного некоторые специализированные клетки обладают специальными органоидами. К ним относятся:
– реснички и жгутики, представляющие собой выросты мембраны клетки, осуществляющие движения клетки. Они имеются у одноклеточных организмов и многоклеточных (кишечный эпителий, сперматозоиды, эпителий дыхательных путей);
– миофибриллы – тонкие нити мышечных клеток, участвующие в сокращении мышц;
– нейрофибриллы – органоиды, характерные для нервных клеток и участвующие в проведении нервных импульсов. Кроме того, в состав клеток входят центриоли – две (иногда более) цилиндрические структуры диаметром около 0,1 мкм и длиной 0,3 мкм. Место расположения центриолей в период между делениями клетки считается серединой клеточного центра. При делении клетки центриоли расходятся в противоположные стороны – к полюсам, определяя ориентацию веретена деления (рис. 16).
Следует иметь в виду, что, хотя животные и растительные клетки имеют много общего, но между ними существуют и серьёзные различия (табл. 3.1).
Более общая классификация клеток представлена на рис. 3.16.
Одно из основных отличий бактерий от архей, состоит в химическом составе мембраны. Бактерии отделены от внешней среды двойным слоем липидов (жиров и жироподобных веществ). Мембраны архей состоят из терпеновых спиртов.
Отличия растительной клетки от животной
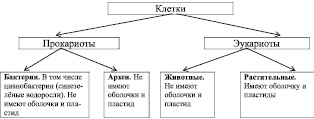
Рис. 3.16. Классификация клеток
При попытке разделить все объекты действительности на живые и неживые выяснилось, что существуют объекты, которые невозможно однозначно отнести ни к живым, ни к неживым. Такие объекты назвали вирусами. Вне клетки они представляют собой неживой химический комплекс ДНК или РНК с молекулами белка. Внутри клетки существование вирусов приобретает форму жизни, то есть они вступают в вещественно-энергетический обмен с содержимым клетки и размножаются.
Таким образом, с учётом живых организмов, классификация всех объектов действительности может выглядеть так, как это представлено на рис. 3.17.
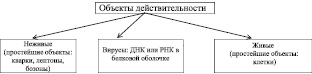
Рис. 3.17. Классификация объектов действительности
1. Обоснуйте утверждение, что ядро – это основная составляющая клетки. Раскройте значение и функции ядра.
2. К какому типу клеток (гаплоидные или диплоидные) относятся следующие примеры: клетка кожи, спора, клетка кожицы лука, клетка печени, сперматозоид, лейкоцит, пыльца, куриное яйцо?
3. В чем состоят основные функции мембранных органоидов: ЭПС, аппарата Гольджи, рибосом и лизосом.
4. Какие существуют виды пластид? Каково их значение для растительной клетки?
5. Что такое митохондрии? Какие процессы в них происходят? Каково их значение?
6. В чем состоят различия между животной и растительной клеткой? Проанализируйте – почему они возникли?
Митохондрии
– микроскопические
двумембранные полуавтономные органоиды
общего назначения, обеспечивающие
клетку энергией, получаемой
благодаря процессам окисления и
запасаемой в виде фосфатных
связей АТФ. Митохондрии
также участвуют в биосинтезе стероидов,
окислении жирных кислот и синтезе
нуклеиновых кислот. Присутствуют во
всех эукариотических клетках. В
прокариотических клетках митохондрий
нет, их функцию выполняют мезосомы –
впячивания наружной цитоплазматической
мембраны внутрь клетки.
Митохондрии могут
иметь эллиптическую, сферическую,
палочковидную, нитевидную и др. формы,
которые могут изменяться в течение
определенного времени. Количество
митохондрий в клетках, выполняющих
различные функции, варьирует в широких
пределах – от 50 и достигая в наиболее
активных клетках 500-5000. Их больше там,
где интенсивны синтетические процессы
(печень) или велики затраты энергии
(мышечные клетки). В клетках печени
(гепатоцитах) их число составляет 800. а
занимаемый ими объем равен примерно
20% объема цитоплазмы. Размеры митохондрий
составляют от 0,2 до 1-2 мкм в диаметре и
от 2 до 5-7 (10) мкм в длину. На светооптическом
уровне митохондрии выявляются в
цитоплазме специальными методами и
имеют вид мелких зерен и нитей (что
обусловило их название – от греч. mitos
– нить и chondros
– зерно).
В цитоплазме
митохондрии могут располагаться
диффузно, однако обычно они сосредоточены
в участках максимального потребления
энергии, например,
вблизи ионных насосов, сократимых
элементов (миофибрилл) органелл движения
(аксонем спермия, ресничек), компонентов
синтетического аппарата (цистерн ЭПС).
Согласно одной из гипотез, все митохондрии
клетки связаны друг с другом и образуют
трехмерную сеть.
Митохондрия
окружена двумя
мембранами – наружной и внутренней,
разделенных
межмембранным
пространством, и
содержат митохондриальный
матрикс, в
который обращены складки внутренней
мембраны – кристы.
Химический состав
внутренней мембраны митохондрий сходен
с мембранами прокариот (например, в ней
присутствует особый липид – кардиодипин
и отсутствует холестерол). Во внутренней
митохондриальной мембране преобладают
белки, составляющие 75%. Во внутреннюю
мембрану встроены белки трех типов (а)
белки
электрон-транспортной цепи (дыхательной
цепи) – НАД’Н-дегидрогеназа и ФАД’Н
дегидрогеназа – и другие транспортные
белки, (б)
грибовидные
тельца АТФ-синтетазы (головки
которых обращены в сторону матрикса)
и (в) часть
ферментов цикла Кребса (сукцинатдегидрогеназа).
Внутренняя
митохондриальная мембрана отличается
чрезвычайно низкой проницаемостью,
транспорт веществ осуществляется через
контактные сайты. Низкая проницаемость
внутренней мембраны для мелких ионов
из-за высокого содержания фосфолипида
Митохондрии
– полуавтономные
органоиды клетки, т.к. содержат собственную
ДНК, полуавтономную систему репликации,
транскрипции и собственный
белоксинтезируюший аппарат – полуавтономную
систему трансляции (рибосомы 70S типа и
т-РНК). Благодаря этому митохондрии
синтезируют часть собственных белков.
Митохондрии могут делиться независимо
от деления клетки. Если из клетки удалить
все митохондрии, то новые в ней не
появятся. Согласно теории эндосимбиоза
митохондрии произошли от аэробных
прокариотических клеток, которые попали
в клетку хозяина, но не переварились,
вступили на путь глубокого симбиоза и
постепенно, утратив автономность,
превратились в митохондрии.
Митохондрии –
полуавтономные
органоиды, что
выражается следующими признаками:
1) наличие
собственного генетического материала
(нити ДНК), что позволяет осуществлять
синтез белка, а также позволяет
самостоятельно делиться независимо
от клетки;
2) наличие двойной
мембраны;
3) пластиды и
митохондрии способны синтезировать
АТФ (для хлоропластов источник энергии
– свет, в митохондриях АТФ образуется
в результате окисления органических
веществ).
1)
Энергетическая
– синтез АТФ (отсюда эти органоиды и
получили название «энергетических
станций клетки»):
При аэробном
дыхание на кристах происходит
окислительное фосфорилирование
(образование АТФ из АДФ и неорганического
фосфата за счет энергии, освободившейся
при окислении органических веществ) и
перенос электронов по электрон-транспортной
цепи. На внутренней мембране митохондрии
расположены ферменты, участвующие в
клеточном дыхании;
2)
участие в биосинтезе
многих соединений (в митохондриях
синтезируются некоторые аминокислоты,
стероиды (стероидогенез), синтезируется
часть собственных белков), а также
накопление ионов (Са2+),
гликопротеидов, белков, липидов;
3) окисление
жирных кислот;
4) генетическая
– синтез
нуклеиновых кислот (идут процессы
репликации и транскрипции). Митохондриальная
ДНК обеспечивает цитоплазматическую
наследственность.
АТФ была открыта
в 1929 году немецким химиком Ломанном. В
1935 году Владимир
Энгельгардт
обратил внимание на то, что мышечные
сокращения невозможны без наличия АТФ.
В период с 1939 под 1941 г. лауреат Нобелевской
премии
Фриц
Липман
доказал, что основным источником энергии
для метаболической реакции является
АТФ, и ввел в обращение термин
“энергетически богатые фосфатные
связи”. Кардинальные изменения в
изучении действия АТФ на организм
произошли в середине 70-х годов, когда
было обнаружено наличие специфических
рецепторов на наружной поверхности
клеточных мембран, чувствительных к
молекуле АТФ. С тех пор интенсивно
изучается триггерное (регуляторное)
действие АТФ на различные функции
организма
Аденозинтрифосфорная
кислота (АТФ,
аденинтрифосфорная кислота) — нуклеотид,
играет исключительно важную роль в
обмене энергии и веществ в организмах;
в первую очередь соединение известно
как универсальный источник энергии
для всех биохимических процессов,
протекающих в живых системах.
Химически АТФ
представляет собой трифосфорный эфир
аденозина, который является производным
аденина
и рибозы.
Пуриновое азотистое
основание
— аденин — соединяется β-N-гликозидной
сзязью с 5′-углеродом рибозы, к которой
последовательно присоединяются три
молекулы фосфорной
кислоты,
обозначаемые соответственно буквами:
α, β и γ.
АТФ относится к
так называемым макроэргическим
соединениям,
то есть к химическим соединениям,
содержащим связи, при гидролизе которых
происходит освобождение значительного
количества энергии. Гидролиз фосфоэфирных
связей молекулы АТФ, сопровождаемый
отщеплением 1 или 2 остатков фосфорной
кислоты, приводит к выделению, по
различным данным, от 40 до 60 кДж/моль.
Высвобождённая
энергия используется в разнообразных
процессах, протекающих с затратой
энергии
1)Главная –
энергетическая. АТФ служит непосредственным
источником энергии для множества
энергозатратных биохимических и
физиологических процессов.
2) синтез нуклеиновых
кислот.
3) регуляция
множества биохимических процессов.
АТФ, присоединяясь к регуляторным
центрам ферментов, усиливает или
подавляет их активность.
В организме АТФ
синтезируется из АДФ,
используя энергию окисляющихся веществ:
Фосфорилирование
АДФ возможно двумя способами: субстратное
фосфорилирование
и окислительное
фосфорилирование.
Основная масса АТФ образуется на
мембранах в митохондриях
путём окислительного фосфорилирования
ферментом H-зависимой
АТФ-синтетазой.
Субстратное фосфорилирование АДФ не
требует участия мембран, оно происходит
в процессе гликолиза
или путём переноса фосфатной группы с
других макроэргических
соединений.
Реакции
фосфорилирования АДФ и последующего
использования АТФ в качестве источника
энергии образуют циклический процесс,
составляющий суть энергетического
обмена.
В организме АТФ
является одним из самых часто обновляемых
веществ. В течение суток одна молекула
АТФ проходит в среднем 2000—3000 циклов
ресинтеза (человеческий организм
синтезирует около 40 кг в день), то есть
запаса АТФ в организме практически не
создаётся, и для нормальной жизнедеятельности
необходимо постоянно синтезировать
новые молекулы АТФ.
Соседние файлы в папке 4,5.Теория
Энергетический обмен, митохондрии
Каждая клетка –
сложная высокоупорядоченная система
и содержимое клетки находится в состоянии
непрерывного активного метаболизма.
Реакции в клетке
подразделяют на две группы:
1) Реакции синтеза
крупных молекулярных структур, которые
протекают с затратой энергии, называются
реакциями синтеза, ассимиляцией или
анаболизмом.
2) Реакции распада
крупных молекул на более мелкие и
простые, протекающие обычно с высвобождением
энергии. Реакции диссимиляции или
катаболизма.
Реакции ассимиляции
формируют так называемый пластический
тип метаболизма. А реакции диссимиляции
– это реакции энергетического обмена.
Все реакции
пластического и энергетического обмена
можно назвать метаболизмом.
Поступающие в
клетку вещества служат строительным
материалом для биосинтеза клеточных
структур и компонентов клетки, а
во-вторых, эти же вещества являются
источником химической энергии.
У всех эукариотических
клеток можно выделить три типа органоидов
энергетического обмена. В растительных
клетках это тилакоидные мембраны
пластиды,
в животных – внутренние мембраны
митохондрий,
а у аэробных бактерий – сопрягающие
мембраны.
На самом деле, пластиды и митохондрии
имеют сопрягающую мембрану.
Для всех сопрягающих
мембран характерны особенности, которые
отличают эту мембрану от других мембран
эукариотических клеток:
1) Сопрягающие
мембраны способны синтезировать
АТФ за счет
внешних ресурсов.
2) Сопрягающие
мембраны несут цепь
переноса электронов.
3) Сопряженные
мембраны имеют специальные «грибовидные»
тельца, в которых находится фермент,
обеспечивающий синтез молекул АТФ с
названием АТФ-синтетаза.
4) Во всех сопрягающих
мембранах содержится повышенная
концентрация белков и низкая концентрация
холестерина.
Преобладающим
фосфолипидом в этих мембранах является
кардиолипин, наличие которого обуславливает
крайне низкую
избирательную способность.
Основой для
связывания двух биохимических процессов,
протекающих на сопрягающей мембране
является мембранный потенциал, который
создается на сопрягающей мембране.
Митохондрии.
Открыл в 1848 году,
Альтман.
Есть почти у всех
эукариотов. Это подвижные органоиды,
могут менять свою морфологию, объединяться,
разъединяться с помощью микротрубочек.
Различные по форме.
Период жизни
митохондрий короток. У человека они
живут 9 суток. Более 10 суток митохондрии
не живут, т.е., высокая степень обновления.
Основная функция
митохондрий – синтез
АТФ, дающий
энергию для всех жизненных процессов.
Митохондрии встречаются практически
у всех эукариотических клеток, кроме
двух видов паразитических амеб.
Митохондрии являются подвижными
пластичными органоидами, они могут
объединяться в гигантские структуры и
наоборот – разрушаться. Совокупность
всех митохондрий в клетке – хондриом.
Размеры варьируют от 1 до 10 мкм. Форма
чрезвычайно разнообразна. Но несмотря
на разнообразие форм и размеров, для
всех митохондрий характерен единый тип
строения. Митохондрия состоит из двух
мембран, наружной и внутренней, меж
которыми находится межмембранное
пространство, которое по сути является
протонным резервуаром. Внутри –
митохондриальный
матрикс.
Наружная мембрана
характеризуется большим количеством
транспортных белков. Эти белки имеют
специфическое название порины.
Они образуют широкие гидрофильные
каналы в бислое липидов, которые
пропускают молекулы с молекулярной
массой до 10000 Дальтон. На наружной
мембране имеются ферменты, которые
превращают липиды в реакционноспособные
соединения. Белков менее 20%. Из физических
свойств можно отметить, что наружная
мембрана может только необратимо
растягиваться.
Внутренняя мембрана
легко сморщивается и растягивается.
Она образует многочисленные складки,
которые называются кристы. Площадь
внутренней мембраны почти в
пять-семь раз больше,
чем наружной. Кристы бывают двух типов
– пластинчатые
в клетках многоклеточных животных и
растений; трубчатые
в тканях синтезирующих ферменты. Кристы
не статистические образования. Расположены
параллельно или перпендикулярно длинной
оси митохондрий.
Степень развития
крист, их количество определяется
функциональной значимостью клетки. На
внутренней мембране очень низок уровень
холестерина. Много особого фосфолипида
кардиолипида. Благодаря этому мембрана
имеет малую
проницаемость.
При суммарном подсчете и вычислении
соотношения белков и липидов получим,
что белков 75%, а липидов 25%.
Межмембранное
пространство или протонный резервуар.
По химическому составу почти идентичен
цитозолю. Содержится только несколько
ферментов для обеспечения реакции
фосфорилирования.
Матрикс –
высококонцентрированная смесь, более
чем из ста компонентов, большинство из
которых – это ферменты цикла Кребса
(карбоновых кислот). Здесь же располагаются
кольцевые молекулы, митохондриальная
ДНК, тРНК, иРНК и митохондриальные
рибосомы с константой седиментации от
55 до 75S
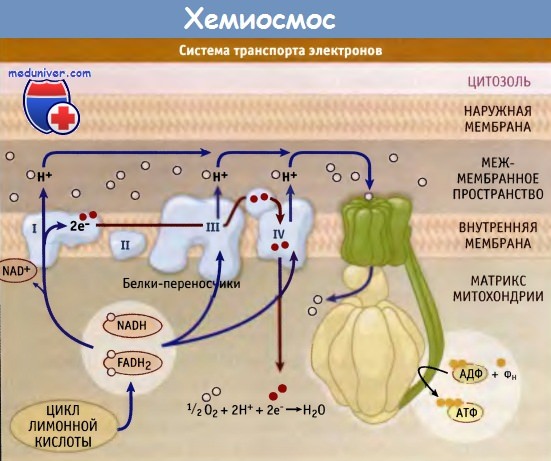
Белки цепи
переноски электронов.
Это крупные ферментные комплексы, по
которым электроны опускаются на все
более низкие энергетические уровни.
Конечным акцептором является молекулярный
кислород. В цепи выделяют электрон-собирающие
и электрон-распределяющие участки и
последовательные ряды ферментов, с
которыми взаимодействуют электроны,
можно разделить на три главных
ферментативных комплекса:
При этом, цитохром
С, принимающий электроны, передает их
на 3 комплекс.
Особую роль играют
Fe содержащие полипептиды. Они различаются
между собой в степени сродства к
электронам, которое выражается через
окислительно-восстановительный потенциал
молекулы. Чем выше этот потенциал, тем
больше сродство окисленной молекулы к
электрону. Электроны переходят по цепи
переноса. Их коферменты восстанавливаются
и окисляются. Цитохромидаза передает
электрон с комплекса цитохрома на
молекулярный кислород. Кислород
активируется и связывается с электронами.
Это важная реакция, в ней кислород
приобретает отрицательный заряд,
благодаря чему водород соединяется с
ним, и образуются вода.
Электроны
перемещаются в толще внутренней мембраны,
при этом через внутреннюю мембрану они
не проходят и остаются на внутренней
стороне мембраны. Протоны обратно не
возвращаются. Они остаются на внешнем
мембранном пространстве. На каждую
исходную молекулы C6H12O6
64 протона. В результате работы дыхательной
цепи на мембранах создается разных
электрических потенциалов. Внутренняя
сторона мембраны заряжается отрицательно
за счет скопления на ней электронов, а
наружная сторона внутренней мембраны
заряжается положительно.
Кроме того, поток
протонов создает между двумя сторонами
внутренней мембраны разность значения
рН. Таким образом, мы имеем кроме
мембранного потенциала еще и градиент
рН. Вместе они составляют электрохимический
протонный градиент. Величина
электрохимического протонного градиента,
который стимулирует синтез, равно 200
мВ. Заствляет ионы двигаться внутрь
матрикса. Заряд – источник протон-движущей
силы.
Грибовидное тело.
Белковый комплекс, который состоит из
двух основных частей: ножки (протон-движущий
канал) и головки (сопрягающего фактора).
Ножка находятся в толще внутренней
мембраны, имеет гидрофобную природу и
по сути является протон-проводящим
канал, по которому протоны из межмембранного
пространства возвращаются в матрикс
митохондрий
Головка тела
называется сопрягающий фактор. Здесь
располагается фермент АТФ-синтетаза,
который превращает энергию движения
протона в энергию органического синтеза,
синтезируя АТФ из молекулы АДФ и
неорганического фосфата (реакция
фосфорилирования).
АТФ занимает 15%
всех белков и образуется при прохождении
1-2 протонов.
Субстратами для
дыхания служат неорганические соединения.
Большинство клеток используют углеводы.
Вовлекаются в процесс расщепления
только после гидролиза до моносахаридов.
Крахмал гидрализуется в глюкозу. Гликоген
гидролизуется примерно до 30000 остатков
глюкоз. Ферментативное расщепление
этих запасных углеводов составляет
первый подготовительный этап
энергетического обмена. Вторым этапом
энергетического обмена является неполное
окисление глюкозы. Оно всегда
бескислородное. Всегда происходит
непосредственно в цитоплазме клетки и
называется гликолизом.
Гликолиз – неполное бескислородное
расщепление глюкозы или многоступенчатый
процесс по превращению углерода в 2
молекулы пировиноградной кислоты
(С3Н4О3).
В ходе гликолиза
выделяется большое количество энергии
(200 кДж на моль), 60% этой энергии рассеивается
в виде тепла, а 40% используется на синтез
АТФ. Кроме двух молекул пирувата,
образуются протоны, которые в цитоплазме
клетки запасаются в форме переносчика.
У животных преобладающим переносчиком
является НАДФ и соединяется с протоном.
Дальнейшая судьба
пирувата имеет три пути:
1) Характерный для
дрожжей и клеток растений при недостатке
кислорода происходит спиртовое брожение
и ПВК восстанавливается до этилового
спирта (С2Н5ОН).
2) В клетках животных
при недостатке кислорода. Накопление
в мышечных волокнах молочной кислоты.
Пируват восстанавливается до молочной
кислоты (С3Н6О3).
3) При наличии в
среде кислорода, продукты гликолиза
расщепляются до СО2
и Н2О
(полное окисление или клеточное дыхание).
Продукты гликолиза переходят для
дальнейшего расщепления в матрикс
митохондрий, где протекает третий этап
энергетического обмена – клеточное
дыхание. Пируват, одновременно с потоком
протонов, направляется внутрь митохондрий.
В матриксе молекулы пирувата взаимодействуют
с каталитическими белками и преобразуется
в молекулу ацетил-коА.
Только он
может вступить в цикл Кребса. Он
вовлекается в цикл трикарбоновых кислот,
который представляет собой цепь
последовательных реакций, в ходе которых
из одной молекулы ацетил-коА образуются
молекулы углекислого газа, 2 АТФ и 4 атома
водорода. Таким образом, в результате
бескислородного окисления и цикла
Кребса, одна молекула глюкозы расщепляется
до диоксида углерода, который легко
выходит из клетки через мембрану и
высвобождающееся при реакции энергия
расходуется в небольшом количестве на
синтез АТФ, а в основном запасается в
нагруженных электронами переносчиков.
Белки-переносчики транспортируют
протоны и электроны к внутренней мембране
митохондрий, где передают их на цепь
дыхательных ферментов.
Транспорт протонов
и электронов приводит к накоплению в
протонном резервуаре протонов. Наружная
поверхность внутренней мембраны заряжена
положительно. Электроны обеспечивают
отрицательный заряд внутренней стороны
мембраны. Когда разность потенциалов
= 200мВ, открывается канал в ножке
грибовидного тела, протоны проталкиваются
через ножку, отдавая энергию ферменты
АТФ-синтетазе, что обеспечивает реакцию
фосфорилирования.
Синтез АТФ сопряжен
с обратным потоком протонов в матрикс.
Протонный градиент
через внутреннюю мембрану митохондрий
дает примерно 24 кДЖ на моль переносимых
протонов. И процесс переноса электронов
(окисление) в мембранах связан с
образованием АТФ из АДФ (реакция
фосфорилирования) и поэтому весь
называется окислительное фосфорилирования.
Ведущую роль в
этих реакциях выполняет внутренняя
сопрягающая мембрана митохондрий. В
процессе окислительного фосфорилирования
перенос каждой пары электронов с
переносчика на молекулярный кислород
доставляет энергию примерно для
образования трех молекул АТФ. При
окислении одной молекулы глюкозы
образуется в реакциях третьего этапа
36 молекул АТФ. Две молекулы АТФ образуются
в бескислородном втором этапе обмена
и поэтому общий выход = 38 молекул АТФ на
одну полностью окисленную молекулу
глюкозы. 55% энергии запасается в виде
химических связей, 45% энергии рассеивается
в виде теплоты.
Сопряжение окисления
и фосфорилирование известно как
хемиосмотическая гипотеза Митчела.
В1978 году он был лауреатом нобелевской
премии за исследование процесса переноса
энергии в клетках и разработку
хемиосмотической теории. Эта теория
объясняет механизм преобразования
энергии в биологической мембране при
синтезе АТФ.
Обмен веществ
между цитоплазмой и матриксом
обеспечивается специальными транспортными
системами, локализованными во внутренней
мембране митохондрии и обеспечивающие
перенос веществ разными механизмами
активного транспорта.
1) По механизма
типа антипорт, когда два вещества
переносятся одновременно в противоположных
направлениях. Большая часть АТФ,
продуцированная митохондриями, выходит
в цитоплазму. Этот перенос осуществляется
АТФ-АДФтранслоказой
2) Симпорт или
сопряженный транспорт. Таким образом,
при участии специфического переносчика
осуществляется одновременный перенос
пирувата и ионов фосфата и протонов.
3) Унипорт. Облегченная
диффузия. Только одно вещество переносится
через мембрану в одном направлении
(ионы кальция внутрь матрикса). С помощью
транспортных белков.
Митохондрия –
полуавтономный органоид. Митохондрии
увеличиваются в количестве при делении
клеток (особенно в период G2),
при увеличении функциональной активности
и нагрузки клетки. Увеличение их
количества происходит путем роста и
деления материнских митохондрий. Делятся
митохондрии путем перетяжки, или
отпочковывания, или возникновения
дочерних митохондрий внутри материнских.
В матриксе
митохондрий содержится собственная
генетическая система, необходимая для
процессов воспроизведения. Это
митохондриальная ДНК, митохондриальные
рибосомы. Гистонов нет. Эти структуры
были обнаружены в 1963 году Нассом.
Митохондриальная ДНК имеет кольцевую
природу. Известно, что все митохондрии
содержат несколько копий своего генома.
От одной копии до десяти. Линейная длина
молекулы варьирует от 5 до 30 мкм. Из
нуклеотидных оснований преимущественно
гуанин и цитозин.
Рибосомы вариабельны,
с константой седиментации 55 – 75S. В 1981
году была расшифрована нуклеотидная
последовательность митохондриального
генома человека. Было показано, что в
митохондриях человека содержится 2 гена
рРНК, 22 гена тРНК и 13 генов, которые
кодируют белки. Репликация и транскрипция
происходят в матриксе митохондрий, и в
этих процессах преобладает контроль
со стороны ядерного генома. Делению или
репродукции митохондрий обязательно
предшествует репликация митохондриальной
ДНК. И этот процесс происходит независимо
от репликации ядерной ДНК. Но обычно
тогда, когда уже ядерная ДНК вся
реплицировалась.
Происхождение и
эволюцию митохондрий объясняют с позиции
эндосимбиотической
гипотезы. И
эта же самая гипотеза представляет
современные митохондрии прямыми
потомками бактериального симбионта.
Это особая группа фотосинтезирующих
бактерий, которые утратили, внедрившись
в организм хозяина, способность к
фотосинтезу, но сохранили дыхательную
функцию. Эти пурпурные бактерии,
внедрившись в другую эукариотическую
клетку, укоренились к ней.
Последние наблюдения
цитологов после расшифровки генома
показали, что митохондрии эволюционировали
вместе со всеми эукариотами от одного
общего предка (коэволюция – совместная
эволюция). Эволюционно, митохондриальный
и ядерные
компоненты
эукариот возникли одновременно.
Митохондрии
– микроскопические
двумембранные полуавтономные органоиды
общего назначения, обеспечивающие
клетку энергией, получаемой
благодаря процессам окисления и
запасаемой в виде фосфатных
связей АТФ. Митохондрии
также участвуют в биосинтезе стероидов,
окислении жирных кислот и синтезе
нуклеиновых кислот. Присутствуют во
всех эукариотических клетках. В
прокариотических клетках митохондрий
нет, их функцию выполняют мезосомы –
впячивания наружной цитоплазматической
мембраны внутрь клетки.
Митохондрии могут
иметь эллиптическую, сферическую,
палочковидную, нитевидную и др. формы,
которые могут изменяться в течение
определенного времени. Количество
митохондрий в клетках, выполняющих
различные функции, варьирует в широких
пределах – от 50 и достигая в наиболее
активных клетках 500-5000. Их больше там,
где интенсивны синтетические процессы
(печень) или велики затраты энергии
(мышечные клетки). В клетках печени
(гепатоцитах) их число составляет 800. а
занимаемый ими объем равен примерно
20% объема цитоплазмы. Размеры митохондрий
составляют от 0,2 до 1-2 мкм в диаметре и
от 2 до 5-7 (10) мкм в длину. На светооптическом
уровне митохондрии выявляются в
цитоплазме специальными методами и
имеют вид мелких зерен и нитей (что
обусловило их название – от греч. mitos
– нить и chondros
– зерно).
В цитоплазме
митохондрии могут располагаться
диффузно, однако обычно они сосредоточены
в участках максимального потребления
энергии, например,
вблизи ионных насосов, сократимых
элементов (миофибрилл) органелл движения
(аксонем спермия, ресничек), компонентов
синтетического аппарата (цистерн ЭПС).
Согласно одной из гипотез, все митохондрии
клетки связаны друг с другом и образуют
трехмерную сеть.
Митохондрия
окружена двумя
мембранами – наружной и внутренней,
разделенных
межмембранным
пространством, и
содержат митохондриальный
матрикс, в
который обращены складки внутренней
мембраны – кристы.
Химический состав
внутренней мембраны митохондрий сходен
с мембранами прокариот (например, в ней
присутствует особый липид – кардиодипин
и отсутствует холестерол). Во внутренней
митохондриальной мембране преобладают
белки, составляющие 75%. Во внутреннюю
мембрану встроены белки трех типов (а)
белки
электрон-транспортной цепи (дыхательной
цепи) – НАД’Н-дегидрогеназа и ФАД’Н
дегидрогеназа – и другие транспортные
белки, (б)
грибовидные
тельца АТФ-синтетазы (головки
которых обращены в сторону матрикса) и
(в) часть
ферментов цикла Кребса (сукцинатдегидрогеназа).
Внутренняя
митохондриальная мембрана отличается
чрезвычайно низкой проницаемостью,
транспорт веществ осуществляется через
контактные сайты. Низкая проницаемость
внутренней мембраны для мелких ионов
из-за высокого содержания фосфолипида
Митохондрии
– полуавтономные
органоиды клетки, т.к. содержат собственную
ДНК, полуавтономную систему репликации,
транскрипции и собственный белоксинтезируюший
аппарат – полуавтономную систему
трансляции (рибосомы 70S типа и т-РНК).
Благодаря этому митохондрии синтезируют
часть собственных белков. Митохондрии
могут делиться независимо от деления
клетки. Если из клетки удалить все
митохондрии, то новые в ней не появятся.
Согласно теории эндосимбиоза митохондрии
произошли от аэробных прокариотических
клеток, которые попали в клетку хозяина,
но не переварились, вступили на путь
глубокого симбиоза и постепенно, утратив
автономность, превратились в митохондрии.
Митохондрии –
полуавтономные
органоиды, что
выражается следующими признаками:
1) наличие
собственного генетического материала
(нити ДНК), что позволяет осуществлять
синтез белка, а также позволяет
самостоятельно делиться независимо от
клетки;
2) наличие двойной
мембраны;
3) пластиды и
митохондрии способны синтезировать
АТФ (для хлоропластов источник энергии
– свет, в митохондриях АТФ образуется в
результате окисления органических
веществ).
1)
Энергетическая
– синтез АТФ (отсюда эти органоиды и
получили название «энергетических
станций клетки»):
При аэробном
дыхание на кристах происходит окислительное
фосфорилирование (образование АТФ из
АДФ и неорганического фосфата за счет
энергии, освободившейся при окислении
органических веществ) и перенос электронов
по электрон-транспортной цепи. На
внутренней мембране митохондрии
расположены ферменты, участвующие в
клеточном дыхании;
2)
участие в биосинтезе
многих соединений (в митохондриях
синтезируются некоторые аминокислоты,
стероиды (стероидогенез), синтезируется
часть собственных белков), а также
накопление ионов (Са2+),
гликопротеидов, белков, липидов;
3) окисление
жирных кислот;
4) генетическая
– синтез
нуклеиновых кислот (идут процессы
репликации и транскрипции). Митохондриальная
ДНК обеспечивает цитоплазматическую
наследственность.
АТФ была открыта
в 1929 году немецким химиком Ломанном. В
1935 году Владимир
Энгельгардт
обратил внимание на то, что мышечные
сокращения невозможны без наличия АТФ.
В период с 1939 под 1941 г. лауреат Нобелевской
премии
Фриц
Липман
доказал, что основным источником энергии
для метаболической реакции является
АТФ, и ввел в обращение термин “энергетически
богатые фосфатные связи”. Кардинальные
изменения в изучении действия АТФ на
организм произошли в середине 70-х годов,
когда было обнаружено наличие специфических
рецепторов на наружной поверхности
клеточных мембран, чувствительных к
молекуле АТФ. С тех пор интенсивно
изучается триггерное (регуляторное)
действие АТФ на различные функции
организма
Аденозинтрифосфорная
кислота (АТФ,
аденинтрифосфорная кислота) — нуклеотид,
играет исключительно важную роль в
обмене энергии и веществ в организмах;
в первую очередь соединение известно
как универсальный источник энергии для
всех биохимических процессов, протекающих
в живых системах.
Химически АТФ
представляет собой трифосфорный эфир
аденозина, который является производным
аденина
и рибозы.
Пуриновое азотистое
основание
— аденин — соединяется β-N-гликозидной
сзязью с 5′-углеродом рибозы, к которой
последовательно присоединяются три
молекулы фосфорной
кислоты,
обозначаемые соответственно буквами:
α, β и γ.
АТФ относится к
так называемым макроэргическим
соединениям,
то есть к химическим соединениям,
содержащим связи, при гидролизе которых
происходит освобождение значительного
количества энергии. Гидролиз фосфоэфирных
связей молекулы АТФ, сопровождаемый
отщеплением 1 или 2 остатков фосфорной
кислоты, приводит к выделению, по
различным данным, от 40 до 60 кДж/моль.
Высвобождённая
энергия используется в разнообразных
процессах, протекающих с затратой
энергии
1)Главная –
энергетическая. АТФ служит непосредственным
источником энергии для множества
энергозатратных биохимических и
физиологических процессов.
2) синтез нуклеиновых
кислот.
3) регуляция
множества биохимических процессов.
АТФ, присоединяясь к регуляторным
центрам ферментов, усиливает или
подавляет их активность.
В организме АТФ
синтезируется из АДФ,
используя энергию окисляющихся веществ:
Фосфорилирование
АДФ возможно двумя способами: субстратное
фосфорилирование
и окислительное
фосфорилирование.
Основная масса АТФ образуется на
мембранах в митохондриях
путём окислительного фосфорилирования
ферментом H-зависимой
АТФ-синтетазой.
Субстратное фосфорилирование АДФ не
требует участия мембран, оно происходит
в процессе гликолиза
или путём переноса фосфатной группы с
других макроэргических
соединений.
Реакции
фосфорилирования АДФ и последующего
использования АТФ в качестве источника
энергии образуют циклический процесс,
составляющий суть энергетического
обмена.
В организме АТФ
является одним из самых часто обновляемых
веществ. В течение суток одна молекула
АТФ проходит в среднем 2000—3000 циклов
ресинтеза (человеческий организм
синтезирует около 40 кг в день), то есть
запаса АТФ в организме практически не
создаётся, и для нормальной жизнедеятельности
необходимо постоянно синтезировать
новые молекулы АТФ.
Механизмы образования АТФ в митохондриях
Все живые клетки обладают способностью превращать энергию окружающей среды в общий интермедиат — АТФ.
Клетка получает энергию за счет пищи, которую она усваивает из окружающей среды. Затем эта энергия должна быть превращена в форму, которую можно распределять по клетке. Общее решение этого вопроса (не только для митохондрий, но и для прокариотических клеток, также превращающих энергию) состоит в запасании энергии в форме универсального вещества, которое можно использовать в любой момент и в любом месте клетки.
Различные типы клеток отличаются деталями энергетической системы, однако общим свойством, характерным для всех клеток, является их способность превращать энергию окружающей среды в АТФ, который представляет собой общую молекулу, при необходимости обеспечивающую энергией отдельные химические реакции.
АТФ образуется двумя путями: в цитозоле и в митохондриях. Первый путь реализуется в цитозоле эукариотических клеток и у бактерий. При этом в результате гликолиза происходит распад глюкозы с образованием пирувата и образуются две молекулы АТФ. Эта реакция может происходить в анаэробных условиях (в отсутствие кислорода).
Второй путь является основным источником создания энергетических запасов и осуществляется в митохондриях эукариотических клеток. Процесс, при котором в митохондриях образуется АТФ, называется окислительным фосфорилированием и осуществляется с участием электрон-транспортной цепи. Пируват, высвобождающийся при гликолизе, поступает в матрикс (люмен) митохондрии, где он распадается и, реагируя с коэнзимом А, образует ацетил-КоА.
После этого, при участии цикла лимонной кислоты с освобождением атомов водорода, ацетильный остаток ацетил-КоА распадается, образуя двуокись углерода. Восстановление переносчика НАД+ в НАДН происходит при участии атомов водорода, и затем при окислении НАДН высвобождаются протон и электрон. Из матрикса ионы водорода (протоны) переносятся через мембрану в межмембранное пространство, а электроны перемещаются вдоль мембраны с участием нескольких транспортных белков. В результате по обе стороны мембраны создается градиент концентрации протонов.
Под действием этого градиента протоны переносятся через мембрану в обратном направлении. Это осуществляется при участии большого белкового комплекса АТФ-синтазы, которая и образует АТФ из АДФ и неорганического фосфата. Этот процесс носит название хемиосмос.
Митохондрии часто называют энергетическими центрами (эукариотических) клеток; они снабжают клетку энергией, необходимой для осуществления метаболических процессов и структурных изменений. Точнее говоря, они превращают энергию, поступающую из внешней среды в формы, которые могут быть использованы клеткой. Постоянство структуры и функции митохондрий у всех клеток эукариот убеждает в том, что эндосимбиоз, в результате которого они возникли, должен был произойти на самом начальном этапе эволюции эукариот.
Видео урок цикл Кребса
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
– Также рекомендуем “Механизмы образования АТФ в хлоропластах”