Вопрос задан 10.06.2023 в 18:01.
Предмет Биология.
Спрашивает Barbq-Junior Андрюха.
АДФ 3)соляная кислота 4)глюкоза
Ответка
Задайте свой вопрос и получите ответ от профессионального преподавателя. Выберите лучший ответ.
5 – 9 классы
Хлоропласты — устройство для сбора солнечной энергии возрастом 3 миллиарда лет. Эта микроскопическая солнечная батарея дает жизнь лесам, полям, планктону морей, а также животным включая нас с вами.

Фотосинтез необходим практически для всего живого, и он является основным источником кислорода в атмосфере.
Фотосинтез окружает нас повсюду. Он происходит под нашими ногами, над нашими головами и в освещенных солнцем зонах водной среды. Но что такое фотосинтез? Почему он так важен? И когда он появился? Ответы на эти и другие вопросы вы найдете ниже.
Фотоси́нтез (от др.-греч. — «свет» и — «соединение», «складывание», «связывание», «синтез») — сложный химический процесс преобразования энергии видимого света (в некоторых случаях инфракрасного излучения) в энергию химических связей органических веществ при участии фотосинтетических пигментов (хлорофилл у растений, бактериохлорофилл у бактерий и бактериородопсин у архей).
Схематическое изображение процесса фотосинтеза, происходящего в растениях
Упрощенное уравнение фотосинтеза, происходящего в растениях
В физиологии растений под фотосинтезом чаще понимается фотоавтотрофная функция — совокупность процессов поглощения, превращения и использования энергии квантов света в различных эндергонических реакциях, в том числе превращения углекислого газа в органические вещества.
Когда начался фотосинтез?
Эволюция фотосинтеза имела огромные последствия для Земли. По мере того как органические вещества фотосинтетической жизни захоранивались в толщах земли, углерод удалялся из атмосферы, позволяя накапливаться кислороду.
Имеющиеся данные свидетельствуют о том, что фотосинтезирующие организмы существовали примерно 3,2-3,5 миллиарда лет назад в виде строматолитов. Строматолиты представляют собой слоистые микробные структуры (обычно чередование светлых и темных слоев), обычно образованные цианобактериями и водорослями, и являются самыми древними известными окаменелостями, а значит, и самым ранним свидетельством жизни на Земле.
Когда этот ранний кислород распространился в верхние слои атмосферы (стратосферу), солнечное излучение превратило молекулы кислорода в озон, в результате чего образовался озоновый слой стратосферы. И, конечно, поскольку озоновый слой поглощает большую часть ультрафиолетового излучения Солнца, он играет важную роль в защите здоровья человека, поэтому маловероятно, что жизнь процветала бы без этого защитного щита.
Что такое фотосинтез?
Фотосинтез — это процесс, в ходе которого синтезируются молекулы углеводов. Он используется растениями, водорослями и некоторыми бактериями для превращения солнечного света, воды и углекислого газа в кислород и энергию в виде сахара. Это, вероятно, самый важный биохимический процесс на планете.
По сути, он забирает углекислый газ, выделяемый всеми дышащими организмами, и возвращает его в атмосферу в виде кислорода.
На скорость фотосинтеза влияют интенсивность освещения, концентрация углекислого газа, поступление воды, температура и наличие минералов. Процесс происходит полностью в хлоропластах, и именно хлорофилл, содержащийся в хлоропластах, придает фотосинтезирующим частям растения зеленый цвет.
Фотосинтез важен и в других частях биосферы. Как морские, так и наземные растения извлекают углекислый газ из атмосферы, и часть его осаждается обратно в виде раковин из карбоната кальция или захоранивается в почве в виде органического вещества.
Без фотосинтеза круговорот углерода не мог бы происходить, и у нас вскоре закончилась бы пища. Со временем атмосфера потеряла бы почти весь газообразный кислород, и большинство организмов исчезло бы.
Экологическая обстановка в мире давно уже перестала радовать земные экосистемы. Множество заводов, без которых человечеству просто не обойтись, выбрасывают ежегодно в атмосферу около 10 миллиардов тон углекислого газа. Многие относятся к этому скептически, утверждая, что количество диоксида углерода не меняется в экосистеме Земли.
На деле, проблема не столько в превышении количества CO2, сколько в нарушении обмена веществ в экосистеме Земли. До начала промышленной деятельности человека углекислый газ, при взаимодействии с водой выпадал в осадок в виде карбонатов, потом переходил в почву, откуда служил для многих растений и водорослей удобрениями. Но это процесс, растянутый на десятки и сотни лет. Человечество же использует запасы миллионов лет в сокращенные сроки, перерабатывая твердые формы углерода в виде нефти и угля. При сжигании этих ископаемых в механизмах и на заводах происходит выброс диоксида углерода в воздух.

Единственный выход это воспользоваться другим механизмом и размножить флору. Фотосинтез — это естественный механизм, предусмотренный природой для переработки CO2. Сегодня эта система нужна, как никогда ранее. Производство диоксида углерода растет и соизмеримо выбросам должно расти количество лесов, джунглей, парков и искусственных насаждений. Растение поглощает углекислый газ и выделяет кислород.
— это процесс разложения органических веществ в живых клетках под действием кислорода. В результате дыхания образуется углекислый газ и вода, а также происходит выделение энергии, необходимой для жизнедеятельности.
Как протекает процесс дыхания
При дыхании растения, как и другие живые существа, из воздуха. А ещё они используют кислород, образующийся в их клетках при фотосинтезе.
Дыхание растений происходит постоянно.
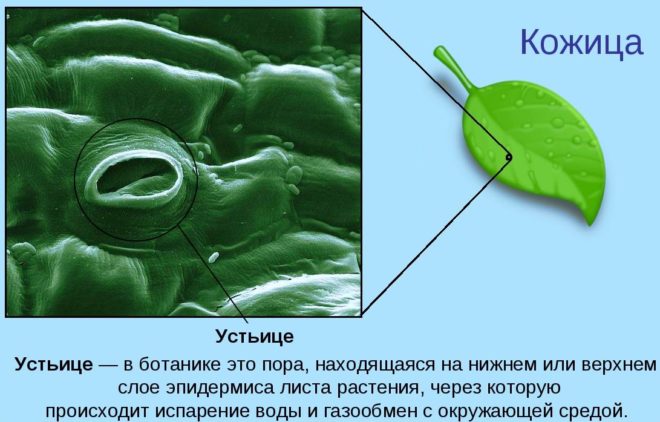
В светлое время суток в эпидермисе листьев и молодых стеблей открыты устьица, кислород из воздуха поступает к клеткам через них.
В тёмное время устьица закрываются. Клетки растений используют кислород, который образовался при фотосинтезе и содержится в межклетниках.
При дыхании происходит распад органических веществ на воду и углекислый газ. А энергия солнечного света, запасённая растениями в ходе фотосинтеза в органических веществах, . Растение может использовать эту энергию на жизненные процессы: на транспорт веществ, размножение, рост и т. д.
Образовавшийся в клетках углекислый газ выходит наружу через устьица и чечевички, или удаляется через оболочки клеток (в корнях).

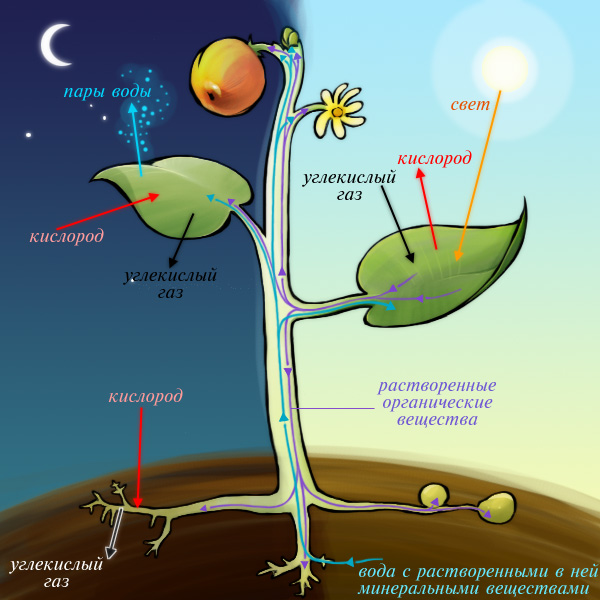
Рис. (1). Процессы фотосинтеза и дыхания
Процесс дыхания противоположен процессу фотосинтеза. Дыхание является обязательным условием существования растений, так как оно обеспечивает растительный организм энергией, необходимой для жизнедеятельности.
Мы уже знаем, что растения корнями впитывают из почвы воду, в которой растворены минеральные соли. Но этого для нормального развития мало. Растениям нужны ещё главные питательные вещества — крахмал и сахар. В почве этих веществ нет, но они есть в растениях.
Учёные установили, что питательные вещества образуются в самих растениях, в их листьях.
Листья растений — это настоящая «кухня», которая может «приготовить» пищу из углекислого газа и воды.
Воду растения получают из почвы благодаря корням. Углекислый газ листья поглощают из воздуха.
Но, чтобы эта волшебная «кухня» заработала, нужен солнечный свет. Солнечный свет даёт энергию, без которой ничего не происходит. Энергия нужна для жизни любого живого существа.
Процесс создания питательных веществ из углекислого газа и воды под действием солнечного света называется .
Фотосинтез происходит в листьях, но в этом процессе принимают участие и другие части растения:
Органические вещества (сахар и крахмал) поступают во все органы растения. Они используются для разных целей:
Учёные сделали ещё одно важное открытие: при фотосинтезе вместе с питательными веществами . Растения выделяют его в воздух.
При фотосинтезе углекислый газ поглощается, а кислород выделяется.
О фотосинтезе уже шла речь в этой статье. Стоит рассмотреть его более подробно. Как уже говорилось ранее, фотосинтез происходит в хлоропластах. За две фазы происходит процесс образования новой молекулы глюкозы, которая после используется в химических процессах растения.
Во время световой фазы используется энергия солнца. Под ее действием вода отдает электрон и распадается на положительно заряженные частицы водорода (Н) и радикалы гидроксида (ОН). После этого оставшиеся частицы ОН образуют воду и кислород, который сразу же удаляется в атмосферу. В хлоропласте остались электроны и положительно заряженные частицы водорода. Эти частицы накапливаются на различных сторонах мембраны тилакоида (одной из частей хлоропластов), из-за разницы концентраций протоны из большей концентрации стремятся проникнуть через мембрану к протонам с меньшей концентрацией. Когда разность потенциалов между ними достигнет 200 миллиВольт, произойдет разряд и молекула АТФ зарядится, а никотинамидадениндинуклеотидфосфат (сокращенно НАДФ) восстановится до НАДФ*Н. Эти два компонента и будут необходимы в темновой фазе фотосинтеза.
Схематический процесс фотосинтеза
В теневой фазе АТФ является аккумулятором, а НАДФ курьером, который доставляет в другую часть хлоропласта протон Н. К тому же растению нужен будет СО2, который послужит основой для будущей молекулы глюкозы. В итоге химических реакций из молекул СО2 и водорода, с помощью энергии из АТФ получается глюкоза С6Н12О6, которая и является первым питательным веществом во всех пищевых цепочках Земли.
Существующие данные указывают на то, что наиболее древними организмами, запасающими энергию света в форме химической энергии, были археи, осуществляющие бесхлорофилльный фотосинтез, при котором не происходит образования восстановителя (НАДФН) и фиксации углекислого газа, а энергия запасается лишь в форме АТФ.
Несколько позже (3,7—3,8 миллиарда лет назад) независимо от архей и многократно в ходе эволюции возникали организмы (зелёные, пурпурные бактерии и другие группы эубактерий) с одной из фотосистем, позволяющей осуществлять аноксигенный фотосинтез, при котором в качестве доноров электронов используются соединения с низким редокс-потенциалом (водород, сульфиды и сероводород, сера, соединения железа (II), нитриты) или осуществляется циклический поток электронов вокруг фотосистемы.
Система оксигенного фотосинтеза, при которой в электрон-транспортной цепи совместно функционируют две фотосистемы, характерная для цианобактерий и происшедших от них в ходе эндосимбиоза хлоропластов водорослей и высших растений, возникла в эволюции однократно, по разным оценкам, от 3,5 до 2,4 миллиардов лет назад. Появление фотосистемы II позволило использовать в качестве неограниченного донора электронов воду — соединение с высоким редокс-потенциалом, то есть не склонное выступать в окислительно-восстановительных процессах в роли восстановителя, однако крайне распространённое на Земле.
С момента появления оксигенных фотосинтезирующих организмов начинается увеличение содержания молекулярного кислорода (сильный окислитель) на Земле, что приводит к накоплению кислорода в водах мирового океана, к окислению горных пород, к формированию озонового экрана и в конечном счёте к накоплению кислорода в атмосфере планеты. Таким образом возникновение системы оксигенного фотосинтеза рассматривается как причина кислородной катастрофы и в частности перестройки древней восстановительной атмосферы Земли и формированию современной атмосферы окислительного типа. Формирование озонового слоя, защищающего поверхность Земли от опасного для живых организмов высокоэнергетического ультрафиолетового излучения, сделало возможным выход жизни на сушу. Одновременно с описанными перестройками гидро-, лито- и атмосферы происходили значительные изменения в биосфере: накопление кислорода привело к смене доминирующих анаэробных сообществ аэробными.
В темновой стадии с участием АТФ и НАДФ происходит восстановление CO2 до глюкозы (C6H12O6). Хотя свет не требуется для осуществления данного процесса, он участвует в его регуляции.
С3-фотосинтез, цикл Кальвина
Цикл Кальвина или восстановительный пентозофосфатный цикл состоит из трёх стадий:
На первой стадии к рибулозо-1,5-бисфосфату присоединяется CO2 под действием фермента рибулозобисфосфат-карбоксилаза/оксигеназа. Этот белок составляет основную фракцию белков хлоропласта и предположительно наиболее распространённый фермент в природе. В результате образуется промежуточное неустойчивое соединение, распадающееся на две молекулы 3-фосфоглицериновой кислоты (ФГК).
Во второй стадии ФГК в два этапа восстанавливается. Сначала она фосфорилируется АТФ под действием фосфороглицерокиназы с образованием 1,3-бисфосфоглицериновой кислоты (ДФГК), затем при воздействии триозофосфатдегидрогеназы и НАДФН ацил-фосфатная группа ДФГК дефосфорилируется и восстанавливается до альдегидной и образуется глицеральдегид-3-фосфат — фосфорилированный углевод (ФГА).
В третьей стадии участвуют 5 молекул ФГА, которые через образование 4-, 5-, 6- и 7-углеродных соединений объединяются в 3 5-углеродных рибулозо-1,5-бифосфата, для чего необходимы 3 АТФ.
Наконец, две ФГА необходимы для синтеза глюкозы. Для образования одной её молекулы требуется 6 оборотов цикла, 6 CO2, 12 НАДФН и 18 АТФ.
При низкой концентрации растворённого в строме CO2 рибулозобифосфаткарбоксилаза катализирует реакцию окисления рибулозо-1,5-бифосфата и его распад на 3-фосфоглицериновую кислоту и фосфогликолевую кислоту, которая вынужденно используется в процессе фотодыхания.
Для увеличения концентрации CO2 растения типа С4 изменили анатомию листа. Цикл Кальвина у них локализуется в клетках обкладки проводящего пучка, в клетках мезофилла же под действием ФЕП-карбоксилазы фосфоенолпируват карбоксилируется с образованием щавелеуксусной кислоты, которая превращается в малат или аспартат и транспортируется в клетки обкладки, где декарбоксилируется с образованием пирувата, возвращаемого в клетки мезофилла.
С4-фотосинтез практически не сопровождается потерями рибулозо-1,5-бифосфата из цикла Кальвина, поэтому более эффективен. Однако он требует не 18, а 30 АТФ на синтез 1 молекулы глюкозы. Это оправдывает себя в тропиках, где жаркий климат требует держать устьица закрытыми, что препятствует поступлению CO2 в лист, а также при рудеральной жизненной стратегии.
Последние заданные вопросы в категории Биология
Биология 26.06.2023 18:34 0 Блындюк Карина.
Биология 26.06.2023 18:33 0 Войтович Арина.
Биология 26.06.2023 18:33 0 Тихомирова Настя.
Биология 26.06.2023 18:01 0 Шмырёва Даша.
Биология 26.06.2023 18:06 0 Лютанов Дима.
Биология 26.06.2023 18:02 0 Сидоренко Дарья.
Биология 26.06.2023 18:02 0 Игорев Игорь.
Биология 26.06.2023 18:46 0 Любушкин Кирилл.
Биология 26.06.2023 18:02 0 Мартинюк Марійка.
- Холл Д., Рао К. Фотосинтез: Пер. с англ. — М.: Мир, 1983.
- Физиология растений / под ред. проф. Ермакова И. П. — М.: Академия, 2007
- Чернавская Н. М., Чернавский Д. С. Туннельный транспорт электронов в фотосинтезе. М., 1977.
- Медведев С. С. Физиология растений — СПб.: СПбГУ, 2006.
Ответы на вопрос
Отвечает Борисенко Ксения.
правильний ответ будет 1) вода
Как происходит фотосинтез?
Растениям требуется световая энергия, углекислый газ, вода и питательные вещества. Эти ингредиенты поступают как из прилегающей атмосферы, так и из почвы.
У живых организмов обнаружено два типа пигментов, способных выполнять функцию фотосинтетических приемников (антенн). Эти пигменты поглощают кванты видимого света и обеспечивают дальнейшее запасание энергии излучения в виде энергии электрохимического градиента ионов H+ (протонов) на биологических мембранах. У подавляющего большинства организмов роль антенн играют хлорофиллы; менее распространён случай, при котором в качестве антенны служит производное витамина А ретиналь. В соответствии с этим выделяют хлорофилльный и бесхлорофилльный фотосинтез.
Система бесхлорофилльного фотосинтеза отличается значительной простотой организации, в связи с чем предполагается эволюционно первичным механизмом запасания энергии электромагнитного излучения. Эффективность бесхлорофилльного фотосинтеза как механизма преобразования энергии сравнительно низка (на один поглощённый квант переносится лишь один H+).
Открытие у галофильных архей
Дитер Остерхельт (Dieter Oesterhelt) и Уолтер Стокениус (Walther Stoeckenius) идентифицировали в «пурпурных мембранах» представителя галофильных архей Halobacterium salinarium (прежнее название Н. halobium) белок, который позже был назван бактериородопсином. Впоследствии были накоплены факты, указывающие на то, что бактериородопсин является светозависимым генератором протонного градиента. В частности, было продемонстрировано фотофосфорилирование на искусственных везикулах, содержащих бактериородопсин и митохондриальную АТФ-синтазу, фотофосфорилирование в интактных клетках H. salinarium, светоиндуцируемое падение pH среды и подавление дыхания, причём все эти эффекты коррелировали со спектром поглощения бактериородопсина. Таким образом, были получены неопровержимые доказательства существования бесхлорофилльного фотосинтеза.
Механизм бесхлорофилльного фотосинтеза галобактерий
Фотосинтетический аппарат экстремальных галобактерий является наиболее примитивным из ныне известных; в нём отсутствует электронтранспортная цепь. Цитоплазматическая мембрана галобактерий является сопрягающей мембраной, содержащей два основных компонента: светозависимый протонный насос (бактериородопсин) и АТФ-синтазу. Работа такого фотосинтетического аппарата основана на следующих трансформациях энергии:
При бесхлорофилльном типе фотосинтеза (как и при реализации циклических потоков в электрон-транспортных цепях) не происходит образования восстановительных эквивалентов (восстановленного ферредоксина или НАД(Ф)Н), необходимых для ассимиляции углекислого газа. Поэтому при бесхлорофилльном фотосинтезе не происходит ассимиляции углекислого газа, а осуществляется исключительно запасание солнечной энергии в форме АТФ (фотофосфорилирование).
Основной путь получения энергии для галобактерий — аэробное окисление органических соединений (при культивировании штаммов используют углеводы и аминокислоты). При дефиците кислорода помимо бесхлорофильного фотосинтеза источниками энергии для галобактерий может служить анаэробное нитратное дыхание или сбраживание аргинина и цитруллина. Однако в эксперименте было показано, что бесхлорофильный фотосинтез может служить и единственным источником энергии в анаэробных условиях при подавлении анаэробного дыхания и брожения при обязательном условии, что в среду вносят ретиналь, для синтеза которого необходим кислород.
Хлорофилльный фотосинтез отличается от бактериородопсинового значительно большей эффективностью запасания энергии. На каждый эффективно поглощённый квант излучения против градиента переносится не менее одного H+, и в некоторых случаях энергия запасается в форме восстановленных соединений (ферредоксин, НАДФ).
Аноксигенный (или бескислородный) фотосинтез протекает без выделения кислорода. К аноксигенному фотосинтезу способны пурпурные и зелёные бактерии, а также гелиобактерии.
При аноксигенном фотосинтезе возможно осуществление:
Оксигенный (или кислородный) фотосинтез сопровождается выделением кислорода в качестве побочного продукта. При оксигенном фотосинтезе осуществляется нециклический электронный транспорт, хотя при определённых физиологических условиях осуществляется исключительно циклический электронный транспорт. В качестве донора электронов при нециклическом потоке используется крайне слабый донор электронов — вода.
Оксигенный фотосинтез распространён гораздо шире. Характерен для высших растений, водорослей, многих протистов и цианобактерий.
Первые опыты по изучению фотосинтеза были проведены Джозефом Пристли в 1770—1780-х годах, когда он обратил внимание на «порчу» воздуха в герметичном сосуде горящей свечой (воздух переставал поддерживать горение, а помещённые в него животные задыхались) и «исправление» его растениями. Пристли сделал вывод, что растения выделяют кислород, который необходим для дыхания и горения, однако не заметил, что для этого растениям нужен свет. Это показал вскоре Ян Ингенхауз.
Позже было установлено, что помимо выделения кислорода растения поглощают углекислый газ и при участии воды синтезируют на свету органическое вещество. В 1842 году Роберт Майер на основании закона сохранения энергии постулировал, что растения преобразуют энергию солнечного света в энергию химических связей. В 1877 году В. Пфеффер назвал этот процесс фотосинтезом.
Хлорофиллы были впервые выделены в 1818 году П. Ж. Пеллетье и Ж. Каванту. Разделить пигменты и изучить их по отдельности удалось М. С. Цвету с помощью созданного им метода хроматографии. Спектры поглощения хлорофилла были изучены К. А. Тимирязевым, он же, развивая положения Майера, показал, что именно поглощённые лучи позволяют повысить энергию системы, создав вместо слабых связей С−О и О−Н высокоэнергетические С−С (до этого считалось, что в фотосинтезе используются жёлтые лучи, не поглощаемые пигментами листа). Сделано это было благодаря созданному им методу учёта фотосинтеза по поглощённому CO2: в ходе экспериментов по освещению растения светом разных длин волн (разного цвета) оказалось, что интенсивность фотосинтеза совпадает со спектром поглощения хлорофилла.
В 1955 году был выделен и очищен фермент рибулозобисфосфат-карбоксилаза/оксигеназа. С4-фотосинтез был описан Ю. С. Карпиловым в 1960 году и М. Д. Хэтчем и Ч. Р. Слэком в 1966 году.
Световая (светозависимая) стадия
В ходе световой стадии фотосинтеза образуются высокоэнергетические продукты: АТФ, служащий в клетке источником энергии, и НАДФ, использующийся как восстановитель. В качестве побочного продукта выделяется кислород.
Фотохимическая суть процесса
Хлорофилл имеет два уровня возбуждения (с этим связано наличие двух максимумов на спектре его поглощения): первый связан с переходом на более высокий энергетический уровень электрона системы сопряжённых двойных связей, второй — с возбуждением неспаренных электронов азота и магния порфиринового ядра. При неизменном спине электрона формируются синглетные первое и второе возбуждённые состояния, при изменённом — триплетное первое и второе.
Второе возбуждённое состояние наиболее высокоэнергетично, нестабильно, и хлорофилл за 10−12 с переходит с него на первое с потерей 100 кДж/моль энергии только в виде теплоты. Из первого синглетного и триплетного состояний молекула может переходить в основное с выделением энергии в виде света (флуоресценция и фосфоресценция соответственно) или тепла с переносом энергии на другую молекулу, либо, поскольку электрон на высоком энергетическом уровне слабо связан с ядром, с переносом электрона на другое соединение.
Первая возможность реализуется в светособирающих комплексах, вторая — в реакционных центрах, где под воздействием кванта света переходящий в возбуждённое состояние хлорофилл становится донором электрона (восстановителем) и передаёт его первичным акцепторам. Чтобы предотвратить возвращение электрона на положительно заряженный хлорофилл, первичный акцептор передаёт его вторичному. Кроме того, время жизни полученных соединений выше, чем у возбуждённой молекулы хлорофилла. Происходит стабилизация энергии и разделение зарядов. Для дальнейшей стабилизации вторичный донор электронов восстанавливает положительно заряженный хлорофилл, первичным же донором в случае оксигенного фотосинтеза является вода.
Хлорофилл выполняет две функции: поглощения и передачи энергии. Более 90 % всего хлорофилла хлоропластов входит в состав светособирающих комплексов (ССК), выполняющих роль антенны, передающей энергию к реакционному центру фотосистем I или II. Помимо хлорофилла, в ССК имеются каротиноиды, а у некоторых водорослей и цианобактерий — фикобилины, роль которых заключается в поглощении света тех длин волн, которые хлорофилл поглощает сравнительно слабо.
Передача энергии идёт резонансным путём (механизм Фёрстера) и занимает для одной пары молекул 10−10—10−12 с, расстояние, на которое осуществляется перенос, составляет около 1 нм. Передача сопровождается некоторыми потерями энергии ( от хлорофилла a к хлорофиллу b, от каротиноидов к хлорофиллу), из-за чего возможна только от пигмента с максимумом поглощения при меньшей длине волны к пигменту с большей. Именно в таком порядке взаимно локализуются пигменты ССК, причём наиболее длинноволновые хлорофиллы находятся в реакционных центрах. Обратный переход энергии невозможен.
ССК растений расположен в мембранах тилакоидов, у цианобактерий основная его часть вынесена за пределы мембран в прикреплённые к ним фикобилисомы — палочковидные полипептидно-пигментные комплексы, в которых находятся различные фикобилины: на периферии фикоэритрины (с максимумом поглощения при ), за ними фикоцианины () и аллофикоцианины (), последовательно передающие энергию на хлорофилл a () реакционного центра.
Основные компоненты цепи переноса электронов
Фотосистема — совокупность ССК, фотохимического реакционного центра и переносчиков электрона. Светособирающий комплекс II содержит 200 молекул хлорофилла a, 100 молекул хлорофилла b, 50 молекул каротиноидов и 2 молекулы феофитина. Реакционный центр фотосистемы II представляет собой пигмент-белковый комплекс, расположенный в тилакоидных мембранах и окружённый ССК. В нём находится димер хлорофилла-a с максимумом поглощения при 680 нм (П680). На него в конечном счёте передаётся энергия кванта света из ССК, в результате чего один из электронов переходит на более высокое энергетическое состояние, связь его с ядром ослабляется, и возбуждённая молекула П680 становится сильным восстановителем (химический потенциал E0 = ).
П680 восстанавливает феофитин, в дальнейшем электрон переносится на хиноны, входящие в состав ФС II и далее на пластохиноны, транспортируемые в восстановленной форме к комплексу b6f. Одна молекула пластохинона переносит 2 электрона и 2 протона, которые берутся из стромы.
Заполнение электронной вакансии в молекуле П680 происходит за счёт воды. В состав ФС II входит водоокисляющий комплекс, содержащий в активном центре 4 иона марганца. Для образования одной молекулы кислорода требуется две молекулы воды, дающие 4 электрона. Поэтому процесс проводится в 4 такта и для его полного осуществления требуется 4 кванта света. Комплекс находится со стороны внутритилакоидного пространства, и полученные 4 протона выбрасываются в него.
Таким образом, суммарный результат работы ФС II — это окисление 2 молекул воды с помощью 4 квантов света с образованием 4 протонов во внутритилакоидном пространстве и 2 восстановленных пластохинонов в мембране.
b6f-комплекс — это насос, перекачивающий протоны из стромы во внутритилакоидное пространство и создающий градиент их концентрации за счёт выделяющейся в окислительно-восстановительных реакциях электронтранспортной цепи энергии. 2 пластохинона дают перекачку 4 протонов. В дальнейшем трансмембранный протонный градиент (pH стромы около 8, внутритилакоидного пространства — 5) используется для синтеза АТФ трансмембранным ферментом АТФ-синтазой.
Светособирающий комплекс I содержит примерно 200 молекул хлорофилла.
В реакционном центре первой фотосистемы находится димер хлорофилла a с максимумом поглощения при 700 нм (П700). После возбуждения квантом света он восстанавливает первичный акцептор — хлорофилл a, тот — вторичный (витамин K1 или филлохинон), после чего электрон передаётся на ферредоксин, который и восстанавливает НАДФ с помощью фермента ферредоксин-НАДФ-редуктазы.
Белок пластоцианин, восстановленный в b6f-комплексе, транспортируется к реакционному центру первой фотосистемы со стороны внутритилакоидного пространства и передаёт электрон на окисленный П700.
Циклический и псевдоциклический транспорт электрона
Помимо полного нециклического пути электрона, описанного выше, обнаружены циклический и псевдоциклический.
Суть циклического пути заключается в том, что ферредоксин вместо НАДФ восстанавливает пластохинон, который переносит его назад на b6f-комплекс. В результате образуется больший протонный градиент и больше АТФ, но не возникает НАДФН.
При псевдоциклическом пути ферредоксин восстанавливает кислород, который в дальнейшем превращается в воду и может быть использован в фотосистеме II. При этом также не образуется НАДФН.
Фаза 2
Сразу под кутикулой и эпидермисом находятся палисадные клетки мезофилла. Эти специализированные клетки имеют вертикальную вытянутую форму и расположены близко друг к другу для максимального поглощения света.
Ниже клеток палисадного мезофилла находится губчатая мезофилловая ткань, которая неплотно упакована для эффективного газообмена. Когда газы движутся внутрь и наружу из этих клеток, они растворяются в тонком слое воды, покрывающем клетки.
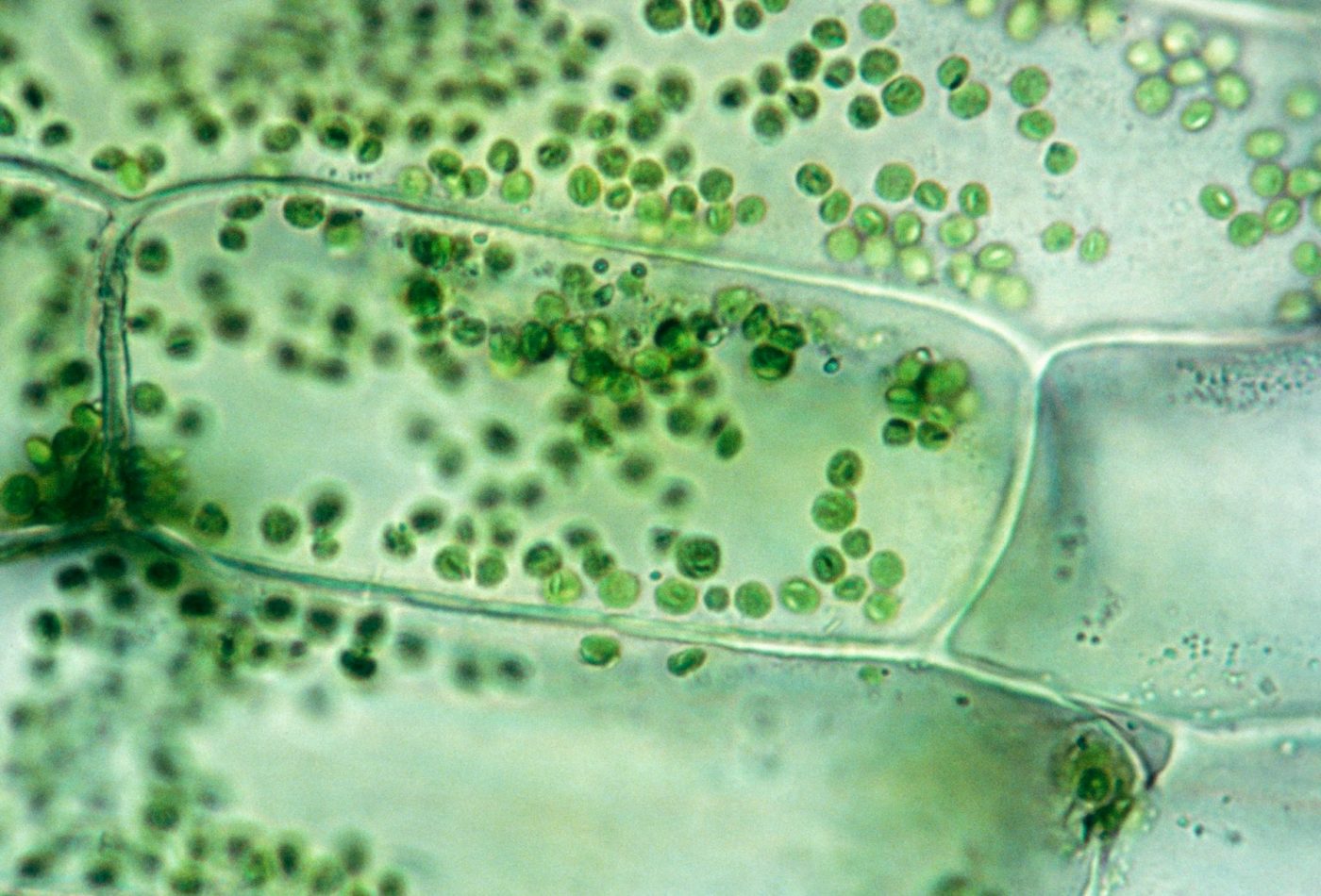
Хлоропласты в клетках водного растения элодея.
Фаза 3
Внутри палисадных клеток мезофилла находятся хлоропласты, много хлоропластов. Они содержат хлорофилл — молекулы, которые не поглощают зеленые волны белого света. Вместо этого они отражают его обратно, придавая растениям зеленый цвет.
Фаза 1
Растения поглощают солнечный свет через два верхних слоя листьев – кутикулу и эпидермис. Эти слои тонкие, поэтому свет легко проходит через них. Углекислый газ поступает из атмосферы, и в то же время вода всасывается из почвы в тело живого растения.
Основная статья: Лист
Фотосинтез растений осуществляется в хлоропластах — полуавтономных двухмембранных органеллах, относящихся к классу пластид. Хлоропласты могут содержаться в клетках стеблей, плодов, чашелистиков, однако основным органом фотосинтеза является лист. Он анатомически приспособлен к поглощению энергии света и ассимиляции углекислоты. Плоская форма листа, обеспечивающая большое отношение поверхности к объёму, позволяет более полно использовать энергию солнечного света. Вода, необходимая для поддержания тургора и протекания фотосинтеза, доставляется к листьям из корневой системы по ксилеме — одной из проводящих тканей растения. Потеря воды в результате испарения через устьица и в меньшей степени через кутикулу (транспирация) служит движущей силой транспорта по сосудам. Однако избыточная транспирация является нежелательной, и у растений в ходе эволюции сформировались различные приспособления, направленные на снижение потерь воды. Отток ассимилятов, необходимый для функционирования цикла Кальвина, осуществляется по флоэме. При интенсивном фотосинтезе углеводы могут полимеризоваться, и при этом в хлоропластах формируются крахмальные зёрна. Газообмен (поступление углекислого газа и выделение кислорода) осуществляется путём диффузии через устьица (некоторая часть газов движется через кутикулу).
Поскольку дефицит углекислого газа значительно увеличивает потери ассимилятов при фотодыхании, необходимо поддерживать высокую концентрацию углекислоты в межклеточном пространстве, что возможно при открытых устьицах. Однако поддержание устьиц в открытом состоянии при высокой температуре приводит к усилению испарения воды, что приводит к водному дефициту и также снижает продуктивность фотосинтеза. Этот конфликт решается в соответствии с принципом адаптивного компромисса. Кроме того, первичное поглощения углекислого газа ночью, при низкой температуре, у растений с CAM-фотосинтезом позволяет избежать высоких транспирационных потерь воды.
У некоторых суккулентов с вырожденными листьями (например, у большинства видов кактусов) основная фотосинтетическая активность связана со стеблем.
Фотосинтез на тканевом уровне
На тканевом уровне фотосинтез у высших растений обеспечивается специализированной тканью — хлоренхимой. Она располагается близ поверхности тела растения, где получает достаточно световой энергии. Обычно хлоренхима находится непосредственно под эпидермой. У растений, растущих в условиях повышенной инсоляции, между эпидермой и хлоренхимой может располагаться один или два слоя прозрачных клеток (гиподерма), обеспечивающих рассеивание света. У некоторых тенелюбивых растений хлоропластами богата и эпидерма (например, кислица). Часто хлоренхима мезофилла листа дифференцирована на палисадную (столбчатую) и губчатую, но может состоять и из однородных клеток. В случае дифференцировки наиболее богата хлоропластами палисадная хлоренхима.
Хлоропласты в клетках листа
Хлоропласты отделены от цитоплазмы двойной мембраной, обладающей избирательной проницаемостью. Внутреннее пространство хлоропласта заполнено бесцветным содержимым (стромой) и пронизано мембранами (ламеллами), которые, соединяясь друг с другом, образуют тилакоиды, которые, в свою очередь, группируются в стопки, называемые гранами. На мембранах тилакоидов располагаются молекулы хлорофилла и других вспомогательных пигментов (каротиноиды). Поэтому их называют фотосинтезирующими мембранами. Внутритилакоидное пространство отделено и не сообщается с остальной стромой; предполагается также, что внутреннее пространство всех тилакоидов сообщается между собой. Световые стадии фотосинтеза приурочены к мембранам, автотрофная фиксация CO2 происходит в строме.
В хлоропластах имеются свои ДНК, РНК, рибосомы (с седиментацией типа 70 S), идёт синтез белка (хотя этот процесс и контролируется из ядра). Они не синтезируются вновь, а образуются путём деления предшествующих. Всё это позволило считать их потомками свободных цианобактерий, вошедших в состав эукариотической клетки в процессе симбиогенеза.
Фотосинтетические мембраны прокариот
Цианобактерии, таким образом, как бы сами являются хлоропластом, и в их клетке фотосинтетический аппарат не вынесен в особую органеллу. Их тилакоиды не образуют стопок, а формируют различные складчатые структуры (у единственной цианобактерии Gloeobacter violaceus тилакоиды отсутствуют вовсе, а весь фотосинтетический аппарат находится в цитоплазматической мембране, не образующей впячиваний). У них и растений также есть различия в светособирающем комплексе (см. ниже) и пигментном составе.
Водоросли и CO2
Под водорослями понимают все растения, находящиеся под водой и не имеющие корня. Интенсивнее всего, из водорослей, поглощает углекислоту одноклеточные водоросли — фитопланктон. В основном все водоросли дышат растворенным в воде кислородом, за исключением нескольких видов, осуществляющих бескислородный фотосинтез. Те в качестве акцептора электронов при дыхании используют элементную серу.
Получение энергии в группе цианобактерий
Фитопланктон обитает в верхних слоях воды, поскольку ему требуется большое количество солнечной энергии для фотосинтеза. При наличии в воде растворенного углекислого газа фитопланктон осуществляет фотосинтезирующий процесс, побочным продуктом которого является кислород. Большим отличием этих водорослей от наземных растений является количество производимого кислорода. За один цикл фотосинтеза фитопланктон производит кислорода в 3-4 раза больше собственного веса. Неудивительно, что при таких показателях 70 процентов атмосферного кислорода произведено в воде.
Фотосинтез — процесс с крайне сложной пространственно-временной организацией.
Разброс характерных времен различных этапов фотосинтеза составляет 19 порядков: скорость процессов поглощения квантов света и миграции энергии измеряется в фемтосекундном интервале (10−15 с), скорость электронного транспорта имеет характерные времена 10−10—10−2 с, а процессы, связанные с ростом растений, измеряются днями (105—107 с).
Также большой разброс размеров характерен для структур, обеспечивающих протекание фотосинтеза: от молекулярного уровня (10−27 м3) до уровня фитоценозов (105 м3).
В фотосинтезе можно выделить отдельные этапы, различающиеся по природе и характерным скоростям процессов:
Какое уравнение фотосинтеза?
Фотосинтезирующие организмы составляют основу пищевой цепи.
Углекислый газ + вода (с энергией света) = глюкоза + кислород
Помимо световой энергии, углекислого газа и воды, растениям необходимы питательные вещества, которые они получают из почвы. Эти питательные вещества снова высвобождаются, или перерабатываются, когда ткани растения отмирают и начинают разлагаться в почве.
Кислород в виде молекул газа (O2) является побочным продуктом фотосинтеза, но именно он отвечает за содержание кислорода в воздухе, который поддерживает нашу жизнь. Растения также выделяют энергию и воду в атмосферу в процессе дыхания.
Сбалансированное уравнение идет немного дальше. Шесть молекул углекислого газа и шесть молекул воды (реагенты) превращаются в одну молекулу сахара (C6H12O6) и шесть молекул кислорода посредством световой энергии, захваченной хлорофиллом.
Фотосинтез составляет энергетическую основу всего живого на планете, кроме хемосинтезирующих бактерий.
Возникновение на Земле более 3 млрд лет назад механизма расщепления молекулы воды квантами солнечного света с образованием O2 представляет собой важнейшее событие в биологической эволюции, сделавшее свет Солнца главным источником энергии биосферы.
Фототрофы обеспечивают конверсию и запасание энергии термоядерных реакций, протекающих на Солнце, в энергию органических молекул. Солнечная энергия при участии фототрофов конвертируется в энергию химических связей органических веществ. Существование гетеротрофных организмов возможно исключительно за счёт энергии, запасённой фототрофами в органических соединениях. При использовании энергии химических связей органических веществ гетеротрофы высвобождают её в процессах дыхания и брожения.
Фотосинтез является основой продуктивности как сельскохозяйственных растений, так и животной пищи.
Энергия, получаемая человечеством при сжигании биотоплива (дрова, пеллеты, биогаз, биодизель, этанол, метанол) и ископаемого топлива (уголь, нефть, природный газ, торф), также является запасённой в процессе фотосинтеза.
Фотосинтез служит главным входом неорганического углерода в биогеохимический цикл.
Большая часть свободного кислорода атмосферы — биогенного происхождения и является побочным продуктом фотосинтеза. Формирование окислительной атмосферы (кислородная катастрофа) полностью изменило состояние земной поверхности, сделало возможным появление дыхания, а в дальнейшем, после образования озонового слоя, позволило жизни существовать на суше.
Топ вопросов за вчера в категории Биология
Биология 14.06.2023 05:14 31 Иваненко Ольга.
Биология 10.06.2023 07:26 11 Топчанский Арсен.
Биология 07.03.2021 17:19 143 Харитонов Кирилл.
Биология 28.02.2021 21:20 31 Федів Антон.
Биология 04.06.2023 01:10 1 Магдеев-Смоляр Тимур.
Биология 24.02.2019 10:05 84 Решетило Артём.
Биология 18.06.2023 16:30 2 Иванов Слава.
Биология 10.06.2023 19:41 5 Козлов Роман.
Биология 05.06.2023 04:25 4 Финапетов Максим.
Фаза 4
Внутри хлоропласта происходит волшебство. Происходит светозависимая реакция, при которой энергия световых волн поглощается и сохраняется в энергонесущих молекулах АТФ.
Затем в светонезависимой реакции (цикл Кальвина) АТФ используется для производства глюкозы, источника энергии. Вода окисляется, углекислый газ восстанавливается, а кислород выбрасывается в атмосферу.
Кислород выделяется через устьица в листьях, микроскопические поры, которые открываются, чтобы впустить углекислый газ и выпустить кислород (и водяной пар).
Фотосинтез и пищевая цепь
Во время фотосинтеза энергия проходит через систему, и вы можете думать о фотосинтезе как о системе потока энергии, прослеживающей путь солнечной энергии через экосистему. Эта энергия запасается первичными продуцентами, фотосинтезирующими организмами. Когда эти организмы поедаются и перевариваются первичными потребителями, высвобождается химическая энергия, которая используется для запуска новых биохимических реакций.
На каждом уровне преобразования энергии по всей пищевой цепи часть энергии теряется в виде тепла. Кроме того, значительная часть энергии, поступающей в каждый организм, используется в процессе дыхания для поддержания жизнедеятельности организма. Эта энергия не сохраняется для использования другими организмами, находящимися выше по пищевой цепи.
Ночное дыхание растений
Процесс дыхания растений мало чем отличается от дыхания животных и человека. Есть и ночное дыхание. Это явление было открыто Отто Варбургом в начале XX века. Ночью света нет, а значит нет и энергии для фотосинтеза. Растения перестают вырабатывать O2, но не могут перестать дышать. Кислород поглощается, а углекислый газ все так же продолжает выделяться.
Белки, жиры и углеводы, запасенные в процессе жизнедеятельности днем, благодаря циклу Кресса превращаются в углекислый газ, молекулы АТФ и водород.
АТФ расходуются на дальнейшие нужды, углекислый газ уходит в атмосферу по устьицам, а вот водород окисляется до воды. Растение не может позволить себе сбрасывать водород в атмосферу, поскольку легко может погибнуть от этого, поэтому происходит частичный выброс паров воды. Большая часть организма растения – вода. Она нужна во всех процессах, включая дневное и ночное дыхание. Окисленный водород будет использован вновь в следующих реакциях.
Именно из-за ночного дыхания не рекомендуется ставить цветы в спальнях. Это увеличивает содержание углекислоты в комнате. Что никак не скажется на цветах, но будет чувствительно для человека.
Для дыхания растений существует пороговое значение содержания кислорода. При увеличении содержания О2 в воздухе до 5-8 процентов – интенсивность дыхания у растений скачкообразно растет. Но после это рост практически прекращается. Сейчас кислорода в воздухе около 21 процента. А значит, растениям еще долго не нужно будет о нем беспокоиться.
В природе есть еще одно интересное явление, названное САМ — фотосинтезом. Это явление характерно для пустынных цветов и растений. В вечной погоне за сохранением водных ресурсов, эти растения приспособились к проведению фотосинтеза в ночь.
Дневное дыхание растений
Дневное дыхание связано с двумя процессами: непосредственно дыханием и фотосинтезом. Процесс дыхания, как и у человека, связан с окислением органических соединений и выделением диоксида углерода, воды и энергии. Вместо человеческих легких выступает вся поверхность растения. Химическая формула, описывающая реакции в процессе
Любое дерево способно дышать всей поверхностью, даже поверхностью плодов. Но наиболее активно процесс дыхания происходит через устья листа, откуда и попадает по межклеточному пространству большая часть необходимых газов.
Если речь идет о дневном времени суток, то дыхание не столь заметно, как ночью. Поскольку работа растения направлена большей частью на постоянное запасание энергии в виде органических соединений (глюкозы). Попадающий в листья газ, при содействии воды и энергии солнечного света в хлоропластах превращается в глюкозу, которую организм запасает для дальнейшего использования. Собственно дыхание и является этим дальнейшим использованием.
Запасенная глюкоза, с помощью воды и кислорода разлагается на молекулы аденозинтрифосфорной кислоты (АТФ), углекислый газ и водород. АТФ – это твердая энергия. Биологический аккумулятор клеток, который обеспечивает энергетическими запасами все живое на планете. Позднее эти запасы будут использованы в жизнедеятельности каждой молекулы организма.
Кажется, что образуется замкнутый круг: фотосинтез происходит с образованием глюкозы и кислорода, но что толку, если потом в результате дыхания растений выделяется диоксид углерода и АТФ. А энергию растения расходуют лично на себя, ничего не оставляя другим. Но весь вопрос в количестве. Далеко не весь кислород, который образуется во время фотосинтеза, поглощается организмом во время дыхания. Растения производят в разы больше, чем поглощают. Может этим они и отличаются от человека. А все энергетические запасы растений рано или поздно переходят в запасы животных или человека. Так растения отдают все свои накопления ради существования экосистемы Земли.
В среднем 1 гектар лесов ежегодно выделяет 4 тонны кислорода и потребляет 5 тонн углекислого газа. Человек в день выдыхает до 1 килограмма диоксида углерода, в год — 365 кг. Следовательно, 1 гектар леса поглощает углекислоту, которую выдыхают 13 человек.
С увеличением процента содержания углекислого газа в атмосфере теоретически можно ускорить рост зеленых насаждений на Земле. Многие исследования показывают, что в условиях теплиц СО2 можно использовать как «воздушное удобрение», ведь иногда при дыхании кислородом растениями поглощается еще и углекислый газ. Но так происходит это только в условиях экспериментов. На открытых пространствах начавшийся рост активизирует насекомых, которые не позволяют лесам и джунглям разрастись. А культурные растения от таких добавок превращаются в легкую добычу для вредителей. Поэтому, чтобы не говорили скептики, нарушение обмена углеродом это плохо.
Разложения углекислого газа
синтеза АТФ
фотолиза воды
восстановления углекислого газа до глюкозы
2. Этот процесс происходит в клетке в ???
3. Источником водорода для восстановления углекислого газа в процессе фотосинтеза является
Ответы на вопрос
Бесплатные вебинары с ответами на все вопросы у нас на канале!