Историческое развитие корня
Филогенетически корень возник позже стебля и листа — в связи с переходом растений к жизни на суше и вероятно, произошёл от корнеподобных подземных веточек. У корня нет ни листьев, ни в определённом порядке расположенных почек. Для него характерен верхушечный рост в длину, боковые разветвления его возникают из внутренних тканей, точка роста покрыта корневым чехликом. Корневая система формируется на протяжении всей жизни растительного организма. Иногда корень может служить местом отложения в запас питательных веществ. В таком случае он видоизменяется.
Виды корней
Главный корень образуется из зародышевого корешка при прорастании семени. От него отходят боковые корни.
Придаточные корни развиваются на стеблях и листьях.
Боковые корни представляют собой ответвления любых корней.
Каждый корень (главный, боковые, придаточные) обладает способностью к ветвлению, что значительно увеличивает поверхность корневой системы, а это способствует лучшему укреплению растения в почве и улучшению его питания.

Типы корневых систем
Различают два основных типа корневых систем: стержневая, имеющая хорошо развитый главный корень, и мочковатая. Мочковатая корневая система состоит из большого числа придаточных корней, одинаковых по величине. Вся масса корней состоит из боковых или придаточных корешков и имеет вид мочки.
Сильно разветвлённая корневая система образует огромную поглощающую поверхность. Например,
Это во много раз превышает площадь надземной массы.

Если у растения хорошо выражен главный корень и развиваются придаточные корни, то формируется корневая система смешанного типа (капуста, помидор).
Внешнее строение корня. Внутреннее строение корня

Корневой чехлик
Корень растёт в длину своей верхушкой, где находятся молодые клетки образовательной ткани. Растущая часть покрыта корневым чехликом, защищающим кончик корня от повреждений, и облегчает продвижение корня в почве во время роста. Последняя функция осуществляется благодаря свойству внешних стенок корневого чехлика покрываться слизью, что уменьшает трение между корнем и частичками почвы. Могут даже раздвигать частички почвы. Клетки корневого чехлика живые, часто содержат зёрна крахмала. Клетки чехлика постоянно обновляются за счёт деления. Участвует в положительных геотропических реакциях (направление роста корня к центру Земли).

Клетки зоны деления активно делятся, протяженность этой зоны у разных видов и у разных корней одного и того же растения неодинакова.
За зоной деления расположена зона растяжения (зона роста). Протяжённость этой зоны не превышает нескольких миллиметров.
По мере завершения линейного роста наступает третий этап формирования корня — его дифференциация, образуется зона дифференциации и специализации клеток (или зона корневых волосков и всасывания). В этой зоне уже различают наружный слой эпиблемы (ризодермы) с корневыми волосками, слой первичной коры и центральный цилиндр.
Строение корневого волоска
Корневые волоски — это сильно удлинённые выросты наружных клеток, покрывающих корень. Количество корневых волосков очень велико (на 1 мм2 от 200 до 300 волосков). Их длина достигает 10 мм. Формируются волоски очень быстро (у молодых сеянцев яблони за 30-40 часов). Корневые волоски недолговечны. Они отмирают через 10-20 дней, а на молодой части корня отрастают новые. Это обеспечивает освоение корнем новых почвенных горизонтов. Корень непрерывно растёт, образуя всё новые и новые участки корневых волосков. Волоски могут не только поглощать готовые растворы веществ, но и способствовать растворению некоторых веществ почвы, а затем всасывать их. Участок корня, где корневые волоски отмерли, некоторое время способен всасывать воду, но затем покрывается пробкой и теряет эту способность.

Оболочка волоска очень тонкая, что облегчает поглощение питательных веществ. Почти всю клетку волоска занимает вакуоль, окружённая тонким слоем цитоплазмы. Ядро находится в верхней части клетки. Вокруг клетки образуется слизистый чехол, который содействует склеиванию корневых волосков с частицами почвы, что улучшает их контакт и повышает гидрофильность системы. Поглощению способствует выделение корневыми волосками кислот (угольной, яблочной, лимонной), которые растворяют минеральные соли.
Корневые волоски играют и механическую роль — они служат опорой верхушке корня, которая проходит между частичками почвы.
Под микроскопом на поперечном срезе корня в зоне всасывания видно его строение на клеточном и тканевом уровнях. На поверхности корня — ризодерма, под ней — кора. Наружный слой коры — экзодерма, вовнутрь от неё — основная паренхима. Её тонкостенные живые клетки выполняют запасающую функцию, проводят растворы питательных веществ в радиальном направлении — от всасывающей ткани к сосудам древесины. В них же происходит синтез ряда жизненно важных для растения органических веществ. Внутренний слой коры — эндодерма. Растворы питательных веществ, поступающие из коры в центральный цилиндр через клетки эндодермы, проходят только через протопласт клеток.

Кора окружает центральный цилиндр корня. Она граничит со слоем клеток, долго сохраняющих способность к делению. Это перицикл. Клетки перицикла дают начало боковым корням, придаточным почкам и вторичным образовательным тканям. Вовнутрь от перицикла, в центре корня, находятся проводящие ткани: луб и древесина. Вместе они образуют радиальный проводящий пучок.
Проводящая система корня проводит воду и минеральные вещества из корня в стебель (восходящий ток) и органические вещества из стебля в корень (нисходящий ток). Состоит она из сосудисто-волокнистых пучков. Основными слагаемыми частями пучка являются участки флоэмы (по ним вещества передвигаются к корню) и ксилемы (по которым вещества передвигаются от корня). Основные проводящие элементы флоэмы — ситовидные трубки, ксилемы — трахеи (сосуды) и трахеиды.
Процессы жизнедеятельности корня
Всасывание воды корневыми волосками из почвенного питательного раствора и проведение её в радиальном направлении по клеткам первичной коры через пропускные клетки в эндодерме к ксилеме радиального проводящего пучка. Интенсивность поглощения воды корневыми волосками называется сосущей силой (S), она равна разнице между осмотическим (P) и тургорным (T) давлением: S=P-T.
Когда осмотическое давление равно тургорному (P=T), то S=0, вода перестаёт поступать в клетку корневого волоска. Если концентрация веществ почвенного питательного раствора будет выше, чем внутри клетки, то вода будет выходить из клеток и наступит плазмолиз — растения завянут. Такое явление наблюдается в условиях сухости почвы, а также при неумеренном внесении минеральных удобрений. Внутри клеток корня сосущая сила корня возрастает от ризодермы по направлению к центральному цилиндру, поэтому вода движется по градиенту концентрации (т. е. из места с большей её концентрацией в место с меньшей концентрацией) и создаёт корневое давление, которое поднимает столбик воды по сосудам ксилемы, образуя восходящий ток. Это можно обнаружить на весенних безлистных стволах, когда собирают «сок», или на срезанных пнях. Истекание воды из древесины, свежих пней, листьев, называется «плачем» растений. Когда распускаются листья, то они тоже создают сосущую силу и притягивают воду к себе — образуется непрерывный столбик воды в каждом сосуде — капиллярное натяжение. Корневое давление является нижним двигателем водного тока, а сосущая сила листьев — верхним. Подтвердить это можно с помощью несложных опытов.
Всасывание воды корнями
Цель: выяснить основную функцию корня.
Что делаем: растение, выращенное на влажных опилках, отряхнём его корневую систему и опустим в стакан с водой его корни. Поверх воды для защиты её от испарения нальём тонкий слой растительного масла и отметим уровень.

Что наблюдаем: через день-два вода в ёмкости опустилась ниже отметки.

Результат: следовательно, корни всосали воду и подали её наверх к листьям.
Можно ещё проделать один опыт, доказывающий всасывание питательных веществ корнем.
Что делаем: срежем у растения стебель оставив пенёк высотой 2-3 см. На пенёк наденем резиновую трубку длиной 3 см, а на верхний конец наденем изогнутую стеклянную трубку высотой 20-25 см.
Что наблюдаем: вода в стеклянной трубке поднимается, и вытекает наружу.
Результат: это доказывает, что воду из почвы корень всасывает в стебель.

А влияет ли температура воды на интенсивность всасывания корнем воды?
Цель: выяснить, как температура влияет на работу корня.
Что делаем: один стакан должен быть с тёплой водой (+17-18ºС), а другой с холодной (+1-2ºС).
Что наблюдаем: в первом случае вода выделяется обильно, во втором — мало, или совсем приостанавливается.
Результат: это является доказательством того, что температура сильно влияет на работу корня.

Тёплая вода активно поглощается корнями. Корневое давление повышается.

Холодная вода плохо поглощается корнями. В этом случае корневое давление падает.
Минеральное питание
Физиологическая роль минеральных веществ очень велика. Они являются основой для синтеза органических соединений, а также факторами, которые изменяют физическое состояние коллоидов, т.е. непосредственно влияют на обмен веществ и строение протопласта; выполняют функцию катализаторов биохимических реакций; воздействуют на тургор клетки и проницаемость протоплазмы; являются центрами электрических и радиоактивных явлений в растительных организмах.
Установлено, что нормальное развитие растений возможно только при наличии в питательном растворе трёх неметаллов — азота, фосфора и серы и — и четырёх металлов — калия, магния, кальция и железа. Каждый из этих элементов имеет индивидуальное значение и не может быть заменён другим. Это макроэлементы, их концентрация в растении составляет 10-2–10%. Для нормального развития растений нужны микроэлементы, концентрация которых в клетке составляет 10-5–10-3%. Это бор, кобальт, медь, цинк, марганец, молибден др. Все эти элементы есть в почве, но иногда в недостаточном количестве. Поэтому в почву вносят минеральные и органические удобрения.
Растение нормально растёт и развивается в том случае, если в окружающей корни среде будут содержаться все необходимые питательные вещества. Такой средой для большинства растений является почва.
Дыхание корней
Для нормального роста и развития растения необходимо чтобы к корню поступал свежий воздух. Проверим, так ли это?
Цель: нужен ли воздух корню?
Что делаем: возьмём два одинаковых сосуда с водой. В каждый сосуд поместим развивающие проростки. Воду в одном из сосудов каждый день насыщаем воздухом с помощью пульверизатора. На поверхность воды во втором сосуде нальём тонкий слой растительного масла, так как оно задерживает поступление воздуха в воду.

Что наблюдаем: через некоторое время растение во втором сосуде перестанет расти, зачахнет, и в конце концов погибнет.

Результат: гибель растения наступает из-за недостатка воздуха, необходимого для дыхания корня.
Видоизменения корней
У некоторых растений в корнях откладываются запасные питательные вещества. В них накапливаются углеводы, минеральные соли, витамины и другие вещества. Такие корни сильно разрастаются в толщину и приобретают необычный внешний вид. В формировании корнеплодов участвуют и корень, и стебель.
Корнеплоды

Если запасные вещества накапливаются в главном корне и в основании стебля главного побега, образуются корнеплоды (морковь). Растения, образующие корнеплоды, в основном двулетники. В первый год жизни они не цветут и накапливают в корнеплодах много питательных веществ. На второй — они быстро зацветают, используя накопленные питательные вещества и образуют плоды и семена.
Корневые клубни

У георгина запасные вещества накапливаются в придаточных корнях, образуя корневые клубни.
Бактериальные клубеньки

Своеобразно изменены боковые корни у клевера, люпина, люцерны. В молодых боковых корешках поселяются бактерии, что способствует усвоению газообразного азота почвенного воздуха. Такие корни приобретают вид клубеньков. Благодаря этим бактериям эти растения способны жить на бедных азотом почвах и делать их более плодородными.
Ходульные

У пандуса, произрастающего в приливно-отливной зоне, развиваются ходульные корни. Они высоко над водой удерживают на зыбком илистом грунте крупные облиственные побеги.
Воздушные

У тропических растений, живущих на ветвях деревьев, развиваются воздушные корни. Они часто встречаются у орхидей, бромелиевых, у некоторых папоротников. Воздушные корни свободно висят в воздухе, не достигая земли и поглощая попадающую на них влагу от дождя или росы.
Втягивающие

У луковичных и клубнелуковичных растений, например у крокусов, среди многочисленных нитевидных корней имеется несколько более толстых, так называемых втягивающих, корней. Сокращаясь, такие корни втягивают клубнелуковицу глубже в почву.
Столбовидные

У фикуса развиваются столбовидные надземные корни, или корни-подпорки.
Почва как среда обитания корней
Почва для растений является средой, из которой оно получает воду и элементы питания. Количество минеральных веществ в почве зависит от специфических особенностей материнской горной породы, деятельности организмов, от жизнедеятельности самих растений, от типа почвы.
Почвенные частицы конкурируют с корнями за влагу, удерживая её своей поверхностью. Это так называемая связанная вода, которая подразделяется на гигроскопическую и плёночную. Удерживается она силами молекулярного притяжения. Доступная растению влага представлена капиллярной водой, которая сосредоточена в мелких порах почвы.
Между влагой и воздушной фазой почвы складываются антагонистические отношения. Чем больше в почве крупных пор, тем лучше газовый режим этих почв, тем меньше влаги удерживает почва. Наиболее благоприятный водно-воздушный режим поддерживается в структурных почвах, где вода и воздух находятся одновременно и не мешают друг другу — вода заполняет капилляры внутри структурных агрегатов, а воздух — крупные поры между ними.
Характер взаимодействия растения и почвы в значительной степени связан с поглотительной способностью почвы — способностью удерживать или связывать химические соединения.
Микрофлора почвы разлагает органические вещества до более простых соединений, участвует в формировании структуры почвы. Характер этих процессов зависит от типа почвы, химического состава растительных остатков, физиологических свойств микроорганизмов и других факторов. В формировании структуры почвы принимают участие почвенные животные: кольчатые черви, личинки насекомых и др.
В результате совокупности биологических и химических процессов в почве образуется сложный комплекс органических веществ, который объединяют термином «гумус».
Метод водных культур
В каких солях нуждается растение, и какое влияние оказывают они на рост и развитие его, было установлено на опыте с водными культурами. Метод водных культур — это выращивание растений не в почве, а в водном растворе минеральных солей. В зависимости от поставленной цели в опыте можно исключить отдельную соль из раствора, уменьшить или увеличить ее содержание. Было выяснено, что удобрения, содержащие азот, способствуют росту растений, содержащие фосфор — скорейшему созреванию плодов, а содержащие калий — быстрейшему оттоку органических веществ от листьев к корням. В связи с этим содержащие азот удобрения рекомендуется вносить перед посевом или в первой половине лета, содержащие фосфор и калий — во второй половине лета.
С помощью метода водных культур удалось установить не только потребность растения в макроэлементах, но и выяснить роль различных микроэлементов.
В настоящее время известны случаи, когда выращивают растения методами гидропоники и аэропоники.
Гидропоника — выращивание растений в сосудах, заполненных гравием. Питательный раствор, содержащий необходимые элементы, подаётся в сосуды снизу.
Аэропоника — это воздушная культура растений. При этом способе корневая система находится в воздухе и автоматически (несколько раз в течение часа) опрыскивается слабым раствором питательных солей.

Разнообразие стеблей
Стебель — осевая часть побега растения, он проводит питательные вещества и выносит листья к свету. В стебле могут откладываться запасные питательные вещества. На нём развиваются листья, цветки, плоды с семенами.
У стебля есть узлы и междоузлия. Узел — участок стебля, на котором находится лист (листья) и почка (почки). Участок стебля между соседними узлами представляет собой междоузлие. Угол, образованный листом и стеблем выше узла, называют листовой пазухой. Почки, занимающие боковое положение на узле, в пазухе листа, называют боковыми или пазушными. На верхушке стебля находится верхушечная почка.
Стебли древесных и травянистых растений отличаются по продолжительности жизни. Надземные побеги трав умеренного климата живу, как правило, один год (продолжительность жизни побегов определяется продолжительностью жизни стебля, листья могут сменяться). У древесных растений стебель существует много лет. Главный стебель дерева называется стволом, у кустарников отдельные крупные стебли называют стволиками.
Существует несколько типов стеблей.
Прямостоячие стебли имеются у многих древесных и травянистых растений (у них рост побегов обычно направлен вверх, к солнцу). Они имеют хорошо развитую механическую ткань, они могут быть одревесневшими (берёза, яблоня) или травянистыми (подсолнечник, кукуруза).

Ползучие стебли стелются по земле и могут укореняться в узлах (живучка ползучая, земляника).

Большое распространение имеют лазающие и вьющиеся стебли, объединяемые в группу лиан. Среди лиан имеются деревянистые и травянистые. Вследствие недостаточного развития арматурных элементов, обусловленного быстротой роста, они нуждаются в опорах. Вьющиеся побеги спирально обвивают опору своими стеблями, причём у одних растений витки спирали направлены по часовой стрелке, а у других — против часовой стрелки. Существуют и нейтральные растения, стебли которых вьются и направо и налево.
Вьющиеся стебли, поднимаясь вверх, обвивают опору (вьюнок полевой, хмель).

Цепляющиеся стебли поднимаются вверх, цепляясь за опору усиками (мышиный горошек, виноград).

Формы стеблей
Если разрезать стебель поперёк, то мы увидим, что на поперечном срезе стебель в очертании чаще всего округлый, с гладким или ребристым краем. Но может быть и другой: трёхгранной (у осоки), четырёхгранной (у крапивы), многогранной (у многих кактусов), сплющенная или плоская (у опунций), крылатая (у душистого горошка).

Широкие плоские стебли, сильно бороздчатые, нередко представляют собой ненормальное разрастание тканей. У злаков стебель (надземная часть) называется соломиной. Он обычно полый в середине (кроме узлов). Полые стебли распространены в семействах зонтичных, тыквенных и др.
Внутреннее строение стебля
Молодые (однолетние) стебли снаружи покрыты кожицей, которая затем замещается пробкой, состоящей из мёртвых клеток, заполненных воздухом. Кожица и пробка — покровные ткани.

Пробка — многослойная покровная ткань. Она появляется уже на первом году жизни побега. С возрастом толщина пробкового слоя увеличивается. Клетки пробки мёртвые, заполнены воздухом, плотно прилегающие друг к другу. Надёжно защищает внутренние ткани стебля от неблагоприятных условий.
Кожица и пробка защищают расположенные глубже клетки стебля от излишнего испарения, различных повреждений, от проникновения внутрь атмосферной пыли с микроорганизмами, вызывающими заболевания растений.
В кожице стебля имеются устьица, через которые происходит газообмен. В пробке развиваются чечевички — маленькие бугорки с отверстиями. Чечевички образованы крупными клетками основной ткани с большими межклетниками.

Кора — под покровной тканью находится кора, внутренняя часть которой представлена лубом. В состав луба, кроме ситовидных трубок и клеток-спутниц, входят клетки, в которых откладываются запасные вещества.
Лубяные волокна, вытянутые клетки с разрушенным содержимым и одревесневшими стенками, представляют механическую ткань стебля. Придают стеблю прочность и повышают сопротивление на изломе.

Ситовидные трубки — это вертикальный ряд вытянутых живых клеток, у которых поперечные стенки пронизаны отверстиями, ядра в этих клетках разрушились, а цитоплазма прилегает к оболочке. Это проводящая ткань луба, по которой перемещаются растворы органических веществ.

Камбий — узкие длинные клетки образовательной ткани с тонкими оболочками. Весной и летом клетки камбия активно делятся — происходит рост стебля в толщину.
Плотный, самый широкий слой — древесина — основная часть стебля. Как и луб, состоит из разных клеток разной формы и величины: сосудами проводящей ткани, древесинными волокнами механической ткани и клетками основной ткани.

Все слои клеток древесины, образовавшиеся весной, летом и осенью, составляют годичное кольцо прироста.
Сердцевина — клетки крупные, тонкостенные, неплотно прилегают друг к другу и выполняют запасающую функцию.
От сердцевины в радиальном направлении через древесину и луб проходят сердцевинные лучи. Они состоят из клеток основной ткани и выполняют запасающую и проводящую функции.
Общие черты анатомического строения стебля
Анатомическое строение стебля соответствует его главным функциям: проводящей — в стебле хорошо развита система проводящих тканей, которая связывает все органы растения; опорной — с помощью механических тканей стебель поддерживает все надземные органы и выносит лист в благоприятные условия освещения; ростовой — в стебле имеется система меристем, поддерживающих нарастание тканей в длину и толщину (верхушечные, боковые, вставочные).
Верхушечная меристема даёт начало первичной боковой меристеме — прокамбию — и вставочным меристемам. В результате деятельности первичных меристем формируется первичная структура стебля. Она может сохраняться у некоторых растений длительное время. Вторичная меристема — камбий — формирует вторичное состояние строения стебля.
Первичная структура. В стебле различают центральный цилиндр (стелу) и первичную кору.
Первичная кора снаружи покрыта эпидермой (покровная ткань), под ней находится хлоренхима (ассимиляционная ткань). Она может образовывать чередующиеся полосы, тянущиеся вдоль стебля, с механическими тканями (колленхимой и склеренхимой).
Центральный цилиндр окружён слоем эндодермы. Основная часть центрального цилиндра занята проводящими тканями (флоэмой и ксилемой), образующими вместе с механической тканью (склеренхимой) сосудисто-волокнистые пучки. Внутрь от проводящих тканей располагается сердцевина, состоящая из неспециализированной паренхимы. Часто в сердцевине образуется воздушная полость.
Вторичная структура — камбий формирует внутрь вторичную ксилему, наружу — вторичную флоэму. Первичная кора отмирает и заменяется вторичной — это совокупность всех вторичных тканей, расположенных снаружи от камбия.
Строение стебля зависит от условий обитания и отражает особенности строения той или иной систематической группы растений.
Внутреннее строение стебля (часть поперечного среза стебля трёхлетнего побега липы)
Перидерма. Первичная покровная ткань (эпидерма) функционирует недолго. Вместо неё образуется вторичная покровная ткань — перидерма, которая состоит из трёх слоёв клеток — пробки (внешний слой), пробкового камбия (средний слой) и феллодермы (внутренний слой). Для осуществления обмена с окружающей средой на перидерме имеются чечевички.
Первичная кора состоит из двух слоёв: колленхимы (слой под перидермой) — механическая ткань — и паренхимы первичной коры (может выполнять запасающую функцию).
Вторичная кора (или луб, флоэма). Типичное строение луба: ситовидные трубки, клетки спутники, лубяная паренхима и лубяные волокна. Лубяные волокна образуют слой, называемый твёрдым лубом; все остальные элементы образуют мягкий луб.
Камбий — образовательная ткань. За счёт деления и дифференциации его клеток снаружи образуются клетки луба (вторичная кора), а внутри — клетки древесины. Как правило, клеток древесины образуется значительно больше, чем клеток коры (соотношение 4:1). Рост стебля в толщину происходит благодаря деятельности клеток камбия. Деятельность камбия прекращается зимой, весной возобновляется.

Древесина (ксилема) — основная часть стебля. Она образуется за счёт деятельности камбия с внутренней его стороны. Состоит из сосудов (трахей), трахеид, древесной паренхимы, древесных волокон (механическая ткань). За год образуется одно кольцо древесины. Граница между годичными кольцами хорошо заметна, потому что весенняя древесина, которая образовалась после пробуждения деятельности камбия, состоит из больших тонкостенных клеток, осенняя — из меньших, более толстостенных клеток. Переход от весенней древесины к осенней постепенный, от осенней к весенней — всегда внезапный (здесь и образуется граница между годичными кольцами). По годичным кольцам древесины можно узнать возраст растения. У тропических растений, которые растут непрерывно в течение года, годичные кольца совсем незаметны.
Сердцевина — центральная часть стебля. Внешний её слой (перимедулярная зона) состоит из живых паренхимных клеток, центральная — из больших клеток, часто отмерших. Между клетками сердцевины могут быть межклеточные пространства. В живых клетках сердцевины откладываются запасные питательные вещества.
Сердцевинный луч — ряд паренхимных клеток, которые начинаются от сердцевины и проходят в радиальном направлении через древесину и луб в первичной коре. Функция их — проводящая и запасающая.
Рост стебля в толщину
Между лубом и древесиной в стебле находится слой клеток камбия. Камбий — это образовательная ткань. Клетки камбия делятся, образуя новые клетки, которые входят в состав древесины и луба. При этом в сторону древесины камбий откладывает клеток больше, чем в сторону коры. Поэтому прирост древесины идёт быстрее, чем луба. В результате деятельности камбия увеличивается толщина стебля.
Условия влияющие на рост дерева в толщину
По толщине годичных колец можно узнать, в каких условиях росло дерево в разные годы жизни. Узкие годичные кольца свидетельствуют о недостатке влаги, о затенении дерева и о плохом питании.

Годичное кольцо — это прирост древесины за год. Во внутренней зоне этого кольца, ближе к сердцевине, сосуды более крупнее и их больше. Это ранняя древесина. В наружной зоне кольца, ближе к коре, клетки более мелкие и более толстостенные. Это — поздняя древесина. Зимой клетки камбия не делятся, они находятся в состоянии покоя. Весной с распусканием почек возобновляется деятельность камбия. Возникают новые клетки древесины и, следовательно, формируется новое годичное кольцо. Крупноклеточная древесина (ранняя) оказывается рядом с мелкоклеточной (поздней) прошлого года. Благодаря такому соседству становится хорошо заметна граница годичными приростами древесины.
Передвижение питательных веществ по стеблю
Для нормальной жизнедеятельности растения вода и питательные вещества должны поступать во все органы. Одна из важнейших функций стебля — транспортная. Она заключается в передаче растворов от органов почвенного питания — корней и органов воздушного питания — листьев ко всем органам растения. В этом легко убедиться, сделав продольный и поперечный срезы стебля растения как показано на рисунке.

Всё растение пронизано проводящими тканями. По одним проводящим тканям движется вода с растворёнными в ней минеральными веществами, по другим — раствор органических веществ. Проводящие ткани объединяются в сосудисто-волокнистые пучки, часто окружённые прочными волокнами механической ткани.
Сосудисто-волокнистые пучки проходят по всему стеблю, соединяя корневую систему с листьями. Но чтобы окончательно убедиться в этом, желательно проделать следующий опыт.
Цель: убедиться, что сосудисто-волокнистые пучки соединяют корневую систему с листьями.
Что делаем: веточку растения поставить на некоторое время в подкрашенную воду. В опыте она заменит минеральные вещества. Через 2-3 часа сделать поперечный и продольный разрез.

Что наблюдаем: изменила свою окраску и стала красной древесина. Кора и сердцевина остались неокрашенными.
Результат: растворы минеральных веществ, как и подкрашенная вода, поднимаются от корня внутри стебля по сосудам древесины. Сосуды проходят через стебель, ответвляются в листья и разветвляются там. По этим сосудам вода с растворёнными в ней минеральными веществами и поступает в листья. Это хорошо видно на продольном и поперечном срезе стебля.
Большое значение для поднятия воды в стебель имеет корневое давление и испарение воды листьями. На место испарившейся воды в листья постоянно поступает новая.
Передвижение по стеблю органических веществ
Органические вещества откладываются в специальных запасающих тканях, из которых одни накапливают эти вещества внутри клеток, другие — внутри клеток и в их оболочках. Вещества, которые откладываются в запас: сахара, крахмал, инулин, аминокислоты, белки, масла.
Органические вещества могут накапливаться в растворённом (в корнеплодах свеклы, чешуйках лука), твёрдом (зёрна крахмала, белка — клубни картофеля, зёрна злаков, бобовых) или полужидком состоянии (капли масла в эндосперме клещевины). Особенно много органических веществ откладывается в видоизменённых подземных побегах (корневищах, клубнях, луковицах), а также в семенах и плодах. В стебле органические вещества могут откладываться в паренхимных клетках первичной коры, сердцевинных лучах, живых клетках сердцевины.
Мы знаем, что крахмал, образовавшийся в листьях, превращается затем в сахар и поступает во все органы растения.
Цель: выяснить, как сахар из листьев проникает в стебель?
Что делаем: на стебле комнатного растения (драцены, фикуса) осторожно сделаем кольцевой надрез. Удалим с поверхности стебля кольцо коры и обнажим древесину. На стебле укрепим стеклянный цилиндр с водой (смотри рисунок).

Что наблюдаем: через несколько недель на ветке, выше кольца появляется утолщение в виде наплыва. На нём начинают развиваться придаточные корни.
Результат: мы знаем, что в лубе расположены ситовидные трубки, а так как, окольцевав ветку мы их перерезали, то органические вещества, оттекающие из листьев, дошли до кольцевой вырезки и скопились там.
Вскоре из наплыва начинают развиваться придаточные корни.

Вывод: таким образом, опыт доказывает, что органические вещества передвигаются по лубу.
Отложение органических веществ
Вода и минеральные соли, всасываемые корнями, передвигаются по стеблю к листьям, цветкам и плодам. Это — восходящий ток, он осуществляется по древесине, основным проводящим элементом которой являются сосуды (мёртвые пустые трубки, образующиеся из живых паренхимных клеток) и трахеиды (мёртвые клетки, которые соединяются между собой с помощью окаймлённых пор).
Органические вещества, образующиеся в листьях, оттекают во все органы растения. Это — нисходящий ток, он осуществляется по лубу, основным проводящим элементом которого являются ситовидные трубки (живые клетки, соединяющиеся между собой ситечками — тонкими перегородками с отверстиями, они могут быть в поперечных и в продольных стенках).
У древесных растений передвижение питательных веществ в горизонтальной плоскости осуществляется с помощью сердцевидных лучей.
Значение запасающей ткани заключается не только в том, что растение при необходимости питается этими органическими веществами, но и в том, что последние являются продуктом питания человека и животных, а также могут использоваться как сырьё.
Физико-механические принципы строения стебля
Тело растения представляет собой систему, которая сильно зависит от воздействия на неё различных метеорологических факторов, а также от давления и веса собственных органов, которые при этом постоянно изменяются в связи с ростом и развитием. Растение постоянно подвергается действию нагрузок как статических, так и динамических. Ему приходится испытывать действие сил ударного характера при различной продолжительности их. К таким силам относятся ветры разной силы и интенсивности, дождь, град, снег и др. надземная часть растения во время ветров, особенно бурь, представляет собой большую парусную поверхность, и легко ломалась бы, если бы не существовали в теле приспособления для сопротивления: прочность — предохраняет от поломки её временными нагрузками. Упругость обеспечивает сопротивление на изгиб, на разрыв. Жёсткость выражается в том, что форма не изменяется существенно от действия механических нагрузок.
Механические ткани играют главную роль в прочности растения. Заякоривание достигается в основании черешков, ветвей и в местах прикрепления корней. Покровная ткань имеет крепкие и утолщённые стенки эпидермиса.
Упругая устойчивость даёт сопротивление при нагрузке сверху на растение. Стебель ветки растения может нагибаться, но не ломаться; например, вертикальные ветки, отягчённые плодами, нагибаются, дают изгиб в виде дуги, но не ломаются, если обладают достаточной упругой устойчивостью. Соломины ржи, пшеницы, ячменя дают дуговые изгибы, если колосья налиты полноценным зерном.
Будучи единым организмом, растение может жить лишь при сочетании этих противоположных принципов (статический — требует распределения тканей на периферии, а сопротивление динамической нагрузки требует распределения материала в центре) распределения тканей прочности.

Первичное строение корня
Первичная структура корня образуется
в результате деятельности апикальной
меристемы. Корень отличается от побега
тем, что его апикальная меристема
откладывает клетки не только внутрь,
но и наружу, пополняя чехлик. Число и
расположение инициальных клеток в
апексах корней значительно варьируют
у растений, принадлежащих к разным
систематическим группам. Производные
инициалей уже вблизи апикальной меристемы
дифференцируются в первичные меристемы:
2) основную меристему
Из этих первичных меристем в зоне
всасывания формируются три системы
тканей:
2) первичная кора
3) осевой (центральный) цилиндр,
или стела.
Ризодерма (эпиблема, эпидерма
корня) – всасывающая ткань, образующаяся
из протодермы, наружного слоя
первичной меристемы корня. В функциональном
отношении ризодерма представляет собой
одну из важнейших тканей растения. Через
нее идет поглощение воды и минеральных
солей, она взаимодействует с живым
населением почвы, через ризодерму из
корня в почву выделяются вещества,
помогающие почвенному питанию. Поглощающая
поверхность ризодермы сильно увеличена
благодаря наличию у части клеток
трубчатых выростов – корневых волосков.
Каждый корневой волосок представляет
собой длинный вырост одной из клеток
ризодермы, в кончике его обычно находится
ядро клетки. Корневой волосок содержит
тонкий пристенный слой цитоплазмы,
более плотный на верхушке волоска, а в
центре – крупную вакуоль. Стенка волоска
очень тонкая и состоит из целлюлозы и
пектиновых веществ. Ее наружные слои
содержат слизь, что способствует
установлению более тесного контакта с
частицами почвы. Слизь создает
благоприятные условия для поселения
полезных бактерий, влияет на доступность
почвенных ионов и защищает корень от
иссушения. В физиологическом отношении
ризодерма отличается большой активностью.
Она поглощает минеральные ионы с затратой
энергии. В гиалоплазме имеется большое
количество рибосом и митохондрий, что
характерно для клеток с высоким уровнем
обмена веществ.
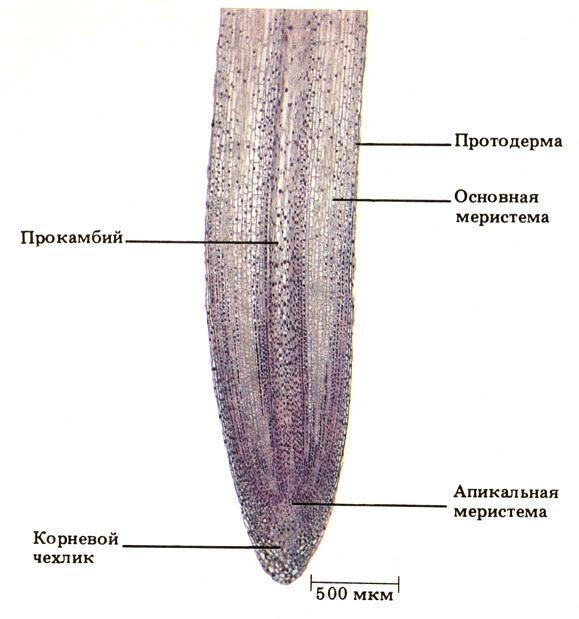
Рис. Продольный срез
кончика корня лука.

Рис. Первичное строение корня
однодольного (а) и
двудольного (б) растений
(поперечные срезы): 1 – центральный
(осевой) цилиндр; 2 – остатки эпиблемы; 3
– экзодерма; 4 – мезодерма; 10 – корневой
волосок
Со временем ризодерма может слущиваться,
и тогда покровную функцию выполняет
экзодерма, а после ее разрушения – слой
клеток мезодермы, иногда мезодермы и
перицикла, стенки которых опробковевают
и одревесневают. Именно поэтому старые
корни однодольных растений имеют меньший
диаметр, чем молодые.
Из основной меристемы формируется
первичная кора. Первичная кора корня
дифференцирована на:
1) экзодерму – наружную часть, лежащую
непосредственно за ризодермой,
2) мезодерму – среднюю часть (паренхима
первичной коры),
3) эндодерму – самый внутренний
слой.
Наружные слои первичной коры, подстилающие
ризодерму, образуют экзодерму.
Экзодерма возникает как ткань, регулирующая
прохождение веществ из ризодермы в
кору, но после отмирания ризодермы выше
зоны всасывания она оказывается на
поверхности корня и превращается в
защитную покровную ткань. Экзодерма
формируется как один слой (реже несколько
слоев) и состоит из живых, многоугольных
по форме, паренхимных клеток, плотно
сомкнутых между собой. Клеточные стенки
пропитаны суберином, т.е. опробковевают.
Это обеспечивает непроницаемость клеток
для воды и газов. В экзодерме, обычно
под корневыми волосками, сохраняются
живые пропускные клетки с тонкими
целлюлозными стенками, через которые
проходят вода и минеральные вещества,
поглощенные ризодермой.
Под экзодермой находятся живые паренхимные
клетки мезодермы. Это
основная масса первичной коры, наиболее
широкая ее часть, образованная живыми
паренхимными клетками с тонкими стенками.
Клетки мезодермы расположены рыхло, по
системе межклетников вдоль оси корня
циркулируют газы, необходимые для
дыхания клеток, выполняют запасающую
функцию, а также функцию проведения
воды и растворенных в ней солей от
корневых волосков в центральный осевой
цилиндр по межклетникам.
У болотных и водных растений, корни
которых испытывают недостаток кислорода,
мезодерма часто представлена аэренхимой.
Также в мезодерме могут присутствовать
механические и выделительные ткани.
Самый внутренний однорядный слой
первичной коры – эндодерма. Она в
виде непрерывного цилиндра окружает
стелу. Клетки эндодермы плотно сомкнуты
и почти квадратные в поперечном сечении.
В зависимости от степени утолщения
клеточной стенки различают два типа
эндодермы:
1) эндодерму с поясками Каспари (на
поперечном срезе они выглядят как пятна
Каспари),
2) эндодерму с подковообразными
утолщениями стенок.
Эндодерма в своем развитии может пройти
три ступени. На первой ступени ее клетки
плотно прилегают друг к другу и имеют
тонкие первичные стенки. На их радиальных
и поперечных стенках образуются утолщения
в виде рамочек – пояски Каспари.
Пояски соседних клеток тесно смыкаются
между собой, так что вокруг стелы
создается их непрерывная система. В
поясках Каспари откладываются суберин
и лигнин, что делает их непроницаемыми
для растворов, т.к. эти вещества закрывают
в местах своего отложения плазмодесменные
канальцы. Поэтому вещества из коры в
стелу и из стелы в кору могут пройти
только по симпласту, т. е. через живые
протопласты клеток эндодермы и под их
контролем. У многих двудольных и
голосеменных растений процесс
дифференциации эндодермы поясками
Каспари заканчивается.
На второй ступени развития суберин
откладывается по всей внутренней
поверхности клеток эндодермы. При этом
некоторые клетки сохраняют первичное
строение. Это пропускные клетки,
они остаются живыми, и через них
осуществляется связь между первичной
корой и центральным цилиндром. Как
правило, они расположены напротив лучей
первичной ксилемы радиального пучка.
У корней, не обладающих вторичным
утолщением, эндодерма может получить
третичное строение. Оно характеризуется
сильным утолщением и одревеснением
всех стенок, или чаще сравнительно
тонкими остаются стенки, обращенные
наружу. Пропускные клетки сохраняются
и в третичной эндодерме. Эндодерма с
подковообразными утолщениями стенок
развивается чаще у однодольных растений,
когда в клетках эндодермы происходит
дальнейшее утолщение клеточных стенок
за счет отложения суберина. Образуется
толстая вторичная клеточная стенка,
которая в дальнейшем одревесневает.
Неутолщенной остается только наружная
клеточная стенка.
Считается, что эндодерма выполняет роль
гидравлического барьера, способствуя
продвижению минеральных веществ и воды
из первичной коры в центральный осевой
цилиндр и препятствуя их выходу обратно.

Рис. Первичное строение корня
ириса (общий план): 1 – экзодерма; 2 – остатки
ризодермы; 3 – паренхима первичной коры;
4 – пропускная клетка эндодермы; 5 –
эндодерма с подковообразными утолщениями;
6 – перицикл; 7 – сосуды ксилемы; 8 – флоэма
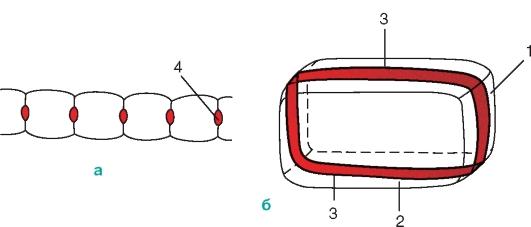
Рис. Схема строения клетки
эндодермы: а – общий вид; б – поперечный
разрез клеток; 1 – поперечная стенка
клетки; 2 – продольная радиальная стенка;
3 – поясок Каспари; 4 – пятна Каспари
Центральный (осевой) цилиндр,
или стела формируется в центре
корня. Уже вплотную к зоне деления самый
наружный слой стелы образует перицикл,
клетки которого долго сохраняют характер
меристемы и способность к новообразованиям.
В молодом корне перицикл состоит из
одного ряда живых паренхимных клеток
с тонкими стенками. Перицикл выполняет
несколько важных функций. У большинства
семенных растений в нем закладываются
боковые корни. У видов с вторичным ростом
он участвует в формировании камбия и
дает начало первому слою феллогена. В
перицикле часто происходит образование
новых клеток, входящих затем в его
состав. У некоторых растений в перицикле
возникают также зачатки придаточных
почек. В старых корнях однодольных
растений клетки перицикла часто
склерифицируются.
За перициклом, в виде центрального тяжа,
находятся клетки прокамбия, которые
дифференцируются в первичные проводящие
ткани. Элементы флоэмы и ксилемы
закладываются по кругу, чередуясь друг
с другом, и развиваются центростремительно.
Однако ксилема в своем развитии обычно
обгоняет флоэму и занимает центр корня.
На поперечном разрезе первичная ксилема
образует звезду, между лучами которой
располагаются участки флоэмы (рис.
4.4). Такая структура получила название
радиального проводящего пучка.
Элементы ксилемы максимально приближены
к поверхности стелы, и в них легче, минуя
флоэму, проникают растворы, поступающие
из коры. Центральная часть корня обычно
занята одним или несколькими крупными
сосудами ксилемы.
Первичная структура корня характерна
для молодых корней всех групп растений.
У споровых и однодольных растений
первичное строение корня сохраняется
в течение всей жизни.
Соседние файлы в папке 6-7 корень
Первичное
строение корня характеризуется наличием
трех систем тканей — покровно-всасывающей
эпидермы с корневыми волосками
(эпиблемы), первичной коры и центрального
цилиндра Эпиблема Выполняет всасывающую
и защитную функции, состоит из одного
слоя тонкостенных, плотно сомкнутых
клеток с корневыми волосками, не
имеет устьиц, толстой кутикулы и трихом.
Первичная кора, Лежащая под элиблемой,
подразделяется на экзодерму, мезодерму
и эндодерму. Экзодерму Составляет 3—4
слоя крупных, плотно сомкнутых клеток
с частично опробковевшими оболочками.
Выполняет защитную и опорно-пропускную
функции. Мезодерма — многослойная
запасающая часть, составляет основную
массу первичной коры. Мезодерма выполняет
также транспортную и воздухоносную
функции. Эндодерма — самый внутренний,
обычно однорядный слой первичной коры,
граничащий с центральным цилиндром,
выполняющий опорно-пропускную функцию.
Центральный, Или Осевой, цилиндр Занимает
срединную часть корня, окружен кольцом
перицикла, из которого формируются
боковые корни, В корне нет настоящей
сердцевины. В центре могут находиться
сосуды, склеренхима, иногда — запасающая
паренхима, образующая Ложную сердцевину
Вторичное строение корня формируется
благодаря появлению и функционированию
вторичных боковых меристем — камбия и
феллогена Камбий образуется из перицикла
над лучами первичной ксилемы и из
прокамбия или паренхимы под флоэмой.
Кнаружи делящийся камбий откладывает
вторичную флоэму, а к центру — вторичную
ксилему. Первичная ксилема оттесняется
к оси корня, а первичная флоэма — к
периферии центрального цилиндра.
Вторичное
строение корня может быть пучкового
типа или беспучкового Пучковый тип
Формируется в том случае, если межпучковый
камбий, образовавшийся над лучами
первичной ксилемы, продуцирует паренхиму
сердцевинных лучей, а пучковый камбий,
лежащий под флоэмой,— вторичную
флоэму и ксилему коллатерального или
биколлатерального проводящего пучка.
При Беспучковом типе Строения кольцо
камбия образует сплошные кольца ксилемы
и флоэмы. Вторичные преобразования в
первичной коре корня связаны с
деятельностью феллогена. Если он
возникает из перицикла, то пробка,
образующаяся кнаружи, изолирует первичную
кору, и она отторгается, слущивается
Если феллоген образуется из паренхимы
мезодермы, что встречается редко, то
часть первичной коры, лежащая под
феллогеном, сохраняется. У древесных
растений корень обычно беспучкового
типа и в древесине формируются кольца
годичного прироста Годичное Кольцо —
это совокупность тканей, образованных
камбием за один вегетационный период.
Покровной тканью корней древесных может
быть перидерма или корка. Наличие в
центре органа радиального проводящего
пучка, склеренхимы или сосудов первичной
ксилемы отличает корень от стебля и
корневища, у которых центральную часть
занимает настоящая сердцевина.
Проводящие ткани. Разнообразие, строение, функции. Проводящие элементы элементы ксилемы и флоэмы. Проводящие пучки.
Проводящие
ткани располагаются в органах растений
в виде продольных тяжей, образуя
проводящие пучки. Различают четыре типа
проводящих пучков: простые, общие,
сложные и сосудисто-волокнистые.Простые
пучки состоят из одного типа проводящих
тканей. Например, в краевых частях
листовых пластинок многих растений
встречаются небольшие по диаметру пучки
из сосудов и трахеид, а в цветоносных
побегах у лилейных – из одних лишь
ситовидных трубок.Общие пучки образуются
трахеидами, сосудами и ситовидными
трубками. Иногда этот термин используется
для обозначения пучков метамера, которые
проходят в междоузлии и являются
листовыми следами. В состав сложных
пучков входят проводящие и паренхимные
ткани. Наиболее совершенными, многообразными
по строению и местоположению являются
сосудисто-волокнистые пучки.
Сосудисто-волокнистые
пучки характерны для многих высших
споровых растений и голосеменных. Однако
они наиболее типичны для покрытосеменных.
В таких пучках выделяются функционально
разные части – флоэма и ксилема. Флоэма
обеспечивает отток ассимилятов из листа
и передвижение их в места использования
или запасания. По ксилеме вода и
растворенные в ней вещества передвигаются
из корневой системы в лист и другие
органы. Объем ксилемной части в несколько
раз превосходит объем флоэмной, поскольку
объем поступающей в растение воды
превышает объем образуемых ассимилятов,
так как значительная часть воды испаряется
растением.
Разнообразие
сосудисто-волокнистых пучков определяется
их происхождением, гистологическим
составом и местонахождением в растении.
Если пучки образуются из прокамбия и
завершают своё развитие по мере
использования запаса клеток образовательной
ткани, как у однодольных, они называются
закрытыми для роста. В отличие от них,
у двудольных открытые пучки не ограничены
в росте, поскольку они формируются
камбием и увеличиваются в диаметре на
протяжении всей жизни растения. В состав
сосудисто-волокнистых пучков кроме
проводящих могут входить основные и
механические ткани. Например, у двудольных
флоэма образуется ситовидными трубками
(проводящая ткань восходящего тока),
лубяной паренхимой (основная ткань) и
лубяными волокнами (механическая ткань).
В состав ксилемы входят сосуды и трахеиды
(проводящая ткань нисходящего тока),
древесинная паренхима (основная ткань)
и древесинные волокна (механическая
ткань). Гистологический состав ксилемы
и флоэмы генетически детерминирован и
может быть использован в систематике
растений для диагностики разных таксонов.
Кроме того, степень развития составных
частей пучков может изменяться под
влиянием условий произрастания растений.
Известно
несколько видов сосудисто-волокнистых
пучков.
Закрытые
коллатеральные проводящие пучки
характерны для листьев и стеблей
однодольных покрытосеменных. В них
отсутствует камбий. Флоэма и ксилема
располагаются бок-о-бок. Для них характерны
некоторые конструктивные особенности.
Так, у пшеницы, отличающейся С3-путём
фотосинтеза, пучки образуются из
прокамбия и имеют первичную флоэму и
первичную ксилему. Во флоэме выделяют
более раннюю протофлоэму и более позднюю
по времени образования, но более
крупноклеточную метафлоэму. Во флоэмной
части отсутствуют лубяная паренхима и
лубяные волокна. В ксилеме первоначально
образуются более мелкие сосуды
протоксилемы, расположенные в одну
линию перпендикулярно к внутренней
границе флоэмы. Метаксилема представлена
двумя крупными сосудами, расположенными
рядом с метафлоэмой перпендикулярно
цепочке сосудов протоксилемы. В этом
случае сосуды располагаются Т-образно.
Известно также V-, Y- и -образное расположение
сосудов. Между сосудами метаксилемы в
1 – 2 ряда расположена мелкоклеточная
склеренхима с утолщенными стенками,
которые по мере развития стебля
пропитываются лигнином. Эта склеренхима
отделяет зону ксилемы от флоэмы. По обе
стороны от сосудов протоксилемы
располагаются клетки древесинной
паренхимы, которые, вероятно, выполняют
трансфузионную роль, поскольку при
переходе пучка из междоузлия в листовую
подушку стеблевого узла они участвуют
в образовании передаточных клеток.
Вокруг проводящего пучка стебля пшеницы
располагается склеренхимная обкладка,
лучше развитая со стороны протоксилемы
и протофлоэмы, около боковых сторон
пучка клетки обкладки располагаются в
один ряд.
У
растений с С4-типом фотосинтеза (кукуруза,
просо и др.) в листьях вокруг закрытых
проводящих пучков располагается обкладка
из крупных клеток хлоренхимы.
Открытые
коллатеральные пучки характерны для
стеблей двудольных. Наличие слоя камбия
между флоэмой и ксилемой, а также
отсутствие склеренхимной обкладки
вокруг пучков обеспечивает их длительный
рост в толщину. В ксилемной и флоэмной
частях таких пучков имеются клетки
основной и механической тканей.
Открытые
коллатеральные пучки могут быть
образованы двумя путями. Во-первых, это
пучки, первично образуемые прокамбием.
Затем в них из клеток основной паренхимы
развивается камбий, производящий
вторичные элементы флоэмы и ксилемы. В
результате пучки будут сочетать
гистологические элементы первичного
и вторичного происхождения. Такие пучки
характерны для многих травянистых
цветковых растений класса Двудольные,
имеющих пучковый тип строения стебля
(бобовые, розоцветные и др.).
Во-вторых,
открытые коллатеральные пучки могут
быть образованы только камбием и состоять
из ксилемы и флоэмы вторичного
происхождения. Они типичны для травянистых
двудольных с переходным типом
анатомического строения стебля (астровые
и др.), а также для корнеплодов типа
свёклы.
В
стеблях растений ряда семейств (Тыквенные,
Пасленовые, Колокольчиковые и др.)
встречаются открытые биколлатеральные
пучки, где ксилема с двух сторон окружена
флоэмой. При этом наружный участок
флоэмы, обращенный к поверхности стебля,
развит лучше внутреннего, а полоска
камбия, как правило, располагается между
ксилемой и наружным участком
флоэмы.Концентрические пучки бывают
двух типов. В амфикрибральных пучках,
характерных для корневищ папоротников,
флоэма окружает ксилему, в амфивазальных
– ксилема кольцом расположена вокруг
флоэмы (корневища ириса, ландыша и др.).
Реже концентрические пучки встречаются
у двудольных (клещевина).
Закрытые
радиальные проводящие пучки образуются
в участках корней, имеющих первичное
анатомическое строение. Радиальный
пучок входит в состав центрального
цилиндра и проходит через середину
корня. Его ксилема имеет вид многолучевой
звезды. Между лучами ксилемы располагаются
клетки флоэмы. Число лучей ксилемы в
значительной мере зависит от генетической
природы растений. Например, у моркови,
свеклы, капусты и других двудольных
ксилема радиального пучка имеет только
два луча. У яблони и груши их может быть
3 – 5, у тыквы и бобов – ксилема четырехлучевая,
а у однодольных – многолучевая. Радиальное
расположение лучей ксилемы имеет
приспособительное значение. Оно сокращает
путь воды от всасывающей поверхности
корня к сосудам центрального цилиндра.
У
многолетних древесных растений и
некоторых травянистых однолетников,
например у льна, проводящие ткани
располагаются в стебле, не образуя четко
выраженных проводящих пучков. Тогда
говорят о непучковом типе строения
стебля.