В любой клетке главным источником энергии для всех протекающих в ней процессов служит универсальное энергетическое соединение — АТФ.

Рис. (1).Строение АТФ
АТФ образуется в результате присоединения к АДФ (аденозиндифосфату) одного остатка фосфорной кислоты. Этот процесс называется :
АДФ + H3PO4+ 40 кДж = АТФ + H2O
В молекуле АТФ есть две богатые энергией химические связи. Это связи между остатками фосфорной кислоты. Такие высокоэнергетические связи называют . При разрыве одной макроэргической связи АТФ превращается в АДФ и выделяется около (40) кДж/моль энергии.
Образование АТФ происходит в процессе энергетического обмена, или диссимиляции.
Энергетический обмен (диссимиляция, катаболизм) — это реакции расщепления и окисления органических веществ, протекающие с выделением энергии, частично расходующейся на образование АТФ.
Энергетический обмен в клетках может быть двухэтапным или трёхэтапным.
В кислородсодержащей среде (у аэробных организмов) диссимиляция протекает в три этапа: подготовительный, бескислородный и кислородный. В результате образуются простые неорганические вещества.

Рис. (2). Этапы энергетического обмена
В бескислородной среде (у анаэробных организмов), а также при недостатке кислорода у аэробных организмов, энергетический обмен протекает в два этапа: подготовительный и бескислородный. В этом случае количество запасённой энергии намного меньше, чем в присутствии кислорода.
Первый этап — подготовительный
На подготовительном этапе большие молекулы органических веществ распадаются до более простых: из полисахаридов образуются моносахариды, из жиров — смесь глицерина и жирных кислот, а из белков — смесь аминокислот.
Этот процесс происходит в лизосомах и в органах пищеварения под действием пищеварительных ферментов.
На подготовительном этапе АТФ не образуется, а небольшое количество выделяющейся энергии рассеивается.
Второй этап — бескислородный (гликолиз)
На втором этапе происходит расщепление продуктов подготовительного этапа под действием ферментов. Кислород при этом не используется.
Бескислородный этап расщепления глюкозы имеет название гликолиз. Этот процесс протекает в цитоплазме клеток.
При гликолизе происходит несколько реакций, в результате которых из молекулы глюкозы
образуется (2) молекулы пировиноградной кислоты (ПВК)
, а также (2) молекулы АТФ. В них запасается около (40) выделившейся энергии, остальные (60) рассеиваются.
Образовавшаяся пировиноградная кислота в анаэробных условиях превращается в молочную кислоту
Такой процесс происходит в клетках животных, некоторых грибов и бактерий. Молочная кислота может образоваться и в мышцах человека при длительных нагрузках и недостатке кислорода. Тогда мышцы начинают болеть.
В растительных клетках и клетках дрожжей из пировиноградной кислоты образуется этиловый спирт
и углекислый газ
, т. к. происходит :
Третий этап — кислородный
В кислородной среде после гликолиза протекает третий этап энергетического обмена — кислородный, или клеточное дыхание. Полное кислородное расщепление осуществляется на мембранах митохондрий.
Этот этап тоже является многостадийным. В нём выделяют два процесса — и .
Сущность третьего (кислородного) этап заключается в окислении ПВК до углекислого газа и воды При этом выделившаяся энергия запасается в (36) молекулах АТФ ((2) — в цикле Кребса и (34) — при окислительном фосфорилировании).
Схема кислородного этапа:
Так как (2) молекулы АТФ образуются при гликолизе, то в сумме при полном окислении одной молекулы глюкозы образуется (38) молекул АТФ.
Суммарное уравнение энергетического обмена:
В реакциях энергетического обмена используется не только глюкоза, но и липиды, белки. Но главным источником энергии в большинстве клеток являются углеводы.
Рис. 1. Строение АТФ. https://image.shutterstock.com/image-illustration/atp-energy-currency-cell-600w-1509423494
СТАДИИ ЭНЕРГЕТИЧЕСКОГО ОБМЕНА
Энергетический обмен (катаболизм, диссимиляция) — это процессы расщепления веществ с высвобождением энергии. Высвобожденная энергия преобразуется в энергию АТФ. Наиболее важными процессами энергетического обмена являются дыхание и брожение.

Энергетический обмен – это совокупность химических реакций постепенного распада органических соединений, сопровождающихся высвобождением энергии, часть которой расходуется на синтез АТФ. Синтезированная АТФ становится универсальным источником энергии для жизнедеятельности организмов. Она образуется в результате реакции фосфорилирования – присоединения остатков фосфорной кислоты к молекуле АДФ. На эту реакцию расходуется энергия, которая затем накапливается в макроэргических связях молекулы АТФ, при распаде молекулы АТФ или при ее гидролизе до АДФ клетка получает около 40 кДж энергии.
АТФ – постоянный источник энергии для клетки, она мобильно может доставлять химическую энергию в любую часть клетки. Когда клетке необходима энергия – достаточно гидролизовать молекулу АТФ. Энергия выделяется в результате реакции диссимиляции (расщепления органических веществ), в зависимости от специфики организма и условий его обитания энергетический обмен проходит в два или три этапа. Большинство живых организмов относятся к аэробам, использующим для обмена веществ кислород, который поступает из окружающей среды. Для аэробов энергетический обмен проходит в три этапа:
В организмах, которые обитают в бескислородной среде и не нуждаются в кислороде для энергетического обмена – анаэробах и аэробах, при недостатке кислорода проходят энергетический обмен в два этапа:
Количество энергии, которое выделяется при двухэтапном варианте намного меньше, чем в трехэтапном.
Подготовительный этап – во время него крупные пищевые полимерные молекулы распадаются на более мелкие фрагменты. В желудочно-кишечном тракте многоклеточных организмов он осуществляется пищеварительными ферментами, у одноклеточных – ферментами лизосом. Полисахариды распадаются на ди- и моносахариды, белки – до аминокислот, жиры – до глицерина и жирных кислот. В ходе этих превращений энергии выделяется мало, она рассеивается в виде тепла, и АТФ не образуется. Образующиеся в ходе подготовительного этапа соединения-мономеры могут участвовать в реакциях пластического обмена (в дальнейшем из них синтезируются вещества, необходимые для клетки) или подвергаться дальнейшему расщеплению с целью получения энергии.
Большинство клеток в первую очередь используют углеводы, жиры остаются в первом резерве и используются по окончания запаса углеводов. Хотя есть и исключения: в клетках скелетных мышц при наличии жирных кислот и глюкозы предпочтение отдается жирным кислотам. Белки расходуются в последнюю очередь, когда запас углеводов и жиров будет исчерпан – при длительном голодании.
Бескислородный этап (гликолиз) – происходит в цитоплазме клеток. Главным источником энергии в клетке является глюкоза. Ее бескислородное расщепление называют анаэробным гликолизом. Он состоит из ряда последовательных реакций по превращению глюкозы в лактат. Его присутствие в мышцах хорошо известно уставшим спортсменам. Этот этап заключается в ферментативном расщеплении органических веществ, полученных в ходе первого этапа. Так как глюкоза является наиболее доступным субстратом для клетки как продукт расщепления полисахаридов, то второй этап можно рассмотреть на примере ее бескислородного расщепления – гликолиза (Рис. 1).

Рис. 1. Бескислородный этап
Гликолиз – многоступенчатый процесс бескислородного расщепления молекулы глюкозы, содержащей шесть атомов углерода, до двух молекул пировиноградной кислоты (пируват). Реакция гликолиза катализируется многими ферментами и протекает в цитоплазме клетки. В ходе гликолиза при расщеплении одного моля глюкозы выделяется около 200 кДж энергии, 60 % ее рассеивается в виде тепла, 40 % – для синтезирования двух молекул АТФ из двух молекул АДФ. При наличии кислорода в среде пировиноградная кислота из цитоплазмы переходит в митохондрии и участвует в третьем этапе энергетического обмена. Если кислорода в клетке нет, то пировиноградная кислота преобразуется в животных клетках или превращается в молочную кислоту.
В микроорганизмах, которые существуют без доступа кислорода – получают энергию в процессе брожения, начальный этап аналогичен гликолизу: распад глюкозы до двух молекул пировиноградной кислоты, и далее она зависит от ферментов, которые находятся в клетке – пировиноградная кислота может преобразовываться в спирт, уксусную кислоту, пропионовую и молочную кислоту. В отличие от того, что происходит в животных тканях, у микроорганизмов этот процесс носит название молочнокислого брожения. Все продукты брожения широко используются в практической деятельности человека: это вино, квас, пиво, спирт, кисломолочные продукты. При брожении, так же как и при гликолизе, выделяется всего две молекулы АТФ.
Кислородный этап стал возможен после накопления в атмосфере достаточного количества молекулярного кислорода, он происходит в митохондриях клеток. Он очень сложен по сравнению с гликолизом, это процесс многостадийный и идет при участии большого количества ферментов. В результате третьего этапа энергетического обмена из двух молекул пировиноградной кислоты формируется углекислый газ, вода и 36 молекул АТФ (Рис. 2).

Рис. 2. Митохондрия
Две молекулы АТФ запасаются в ходе бескислородного расщепления молекулами глюкозы, поэтому суммарный энергетический обмен в клетке в случае распада глюкозы можно представить как:
С6Н12О6 + 6О2 + 38АДФ + 38Н3РО4 = 6СО2 + 44Н2О + 38АТФ
В результате окисления одной молекулы глюкозы шестью молекулами кислорода образуется шесть молекул углекислого газа и выделяется тридцать восемь молекул АТФ.
Мы видим, что в трехэтапном варианте энергетического обмена выделяется гораздо больше энергии, чем в двухэтапном варианте – 38 молекул АТФ против 2.



В отсутствие кислорода или при его недостатке про исходит брожение. Брожение является эволюционно бо лее ранним способом получения энергии, чем дыхание, однако оно энергетически менее выгодно, поскольку конечными продуктами брожения являются органические вещества, богатые энергией. Существует несколько видов брожения, названия которых определяются конечными продуктами: молочнокислое, спиртовое, уксуснокислое и др. Так, в скелетных мышцах в отсутствие кислорода протекает молочнокислое брожение, в ходе которого пировиноградная кислота восстанавливается до молочной кислоты. При этом восстановленные ранее коферменты НАДН расходуются на восстановление пирувата:

Энергетическая эффективность молочнокислого брожения составляет две молекулыАТФ, образованные в процессе окисления глюкозы до пирувата.
Для многих микроорганизмов брожение является единственным способом ассимиляции энергии. Большинство таких организмов живет в анаэробных условиях и погибает в присутствии кислорода, но есть и такие, которые нормально существуют и в присутствии кислорода, и без него. Например, дрожжевые грибы при спиртовом брожении окисляют пировиноградную кислоту до этилового спирта и оксида углерода (IV):


Анаэробный катаболизм глюкозы. Последовательность реакций.
Аэробное окисление глюкозы включает реакции гликолиза и последующее окисление пирувата в цикле Кребса и дыхательной цепи до СО2 и Н2О.
При аэробных условиях пируват проникает в митохондрии, где полностью окисляется до СО2 и Н2О. Если содержание кислороданедостаточно, как это может иметь место в активно сокращающейся мышце, пируват превращается в лактат.
В процессе гликолиза образуется АТФ. Суммарное уравнение гликолиза можно представить следующим образом:

Анаэробный распад включает те же реакции специфического пути распада глюкозы до пирувата, но с последующим превращением пирувата в лактат.
Анаэробный гликолиз отличается от аэробного только наличием последней 11 реакции, первые 10 реакций у них общие.
Лактат не является конечным продуктом метаболизма, удаляемым из организма. Из анаэробной ткани лактат переноситься кровью в печень, где превращаясь в глюкозу (Цикл Кори), или в аэробные ткани (миокард), где превращается в ПВК и окисляется до СО2 и Н2О.
При аэробных условиях пируват проникает в митохондрии, где полностью окисляется до СО2 и Н2О. Если содержание кислорода недостаточно, как это может иметь место в активно сокращающейся мышце, пируват превращается в лактат.
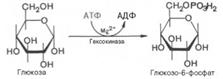

Второй реакцией гликолиза (Аэробное окисление глюкозы) является превращение глюкозо-6-фос-фата под действием фермента глюкозо-6-фосфат-изомеразы во фруктозо-6-фосфат: Эта реакция протекает легко в обоих направлениях, и для нее не требуется каких-либо кофакторов.
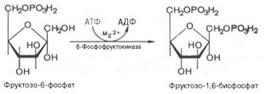
Третья реакция катализируется ферментом фосфофруктокиназой; образовавшийся фруктозо-6-фосфат вновь фосфорилируется за счет второй молекулы АТФ:
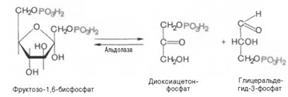

Пятая реакция – это реакция изомеризации триозофосфатов. Катализируется ферментом триозофосфатизомеразой:
Образованием глицеральдегид-3-фосфата как бы завершается первая стадия гликолиза. Вторая стадия – наиболее сложная и важная. Она включает окислительно-восстановительную реакцию (реакция гликолитической оксидоредукции), сопряженную с субстратным фосфорилированием, в процессе которого образуется АТФ.
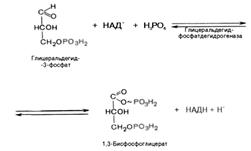
В результате шестой реакции глицеральдегид-3-фосфат в присутствии фермента глицеральдегидфосфатдегидрогеназы, кофермента НАД и неорганического фосфата подвергается своеобразному окислению с образованием 1,3-бисфосфоглицериновой кислоты и восстановленной формы НАД (НАДН). Эта реакция блокируется йод- или бромацетатом, протекает в несколько этапов:
1,3-Бисфосфоглицерат представляет собой высокоэнергетическое соединение (макроэргическая связь условно обозначена знаком «тильда» ~).


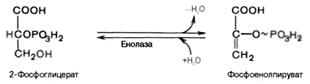
Восьмая реакция сопровождается внутримолекулярным переносом оставшейся фосфатной группы, и 3-фосфоглицериновая кислотапревращается в 2-фосфоглицериновую кислоту (2-фосфоглицерат).
Девятая реакция катализируется ферментом енолазой, при этом 2-фосфоглицериновая кислота в результате отщепления молекулыводы переходит в фосфоенолпировиноградную кислоту (фосфоенолпируват), а фосфатная связь в положении 2 становится высокоэргической:
Десятая реакция характеризуется разрывом высокоэргической связи и переносом фосфатного остатка от фосфоенолпирувата на АДФ(субстратное фосфорилирование). Катализируется ферментом пируваткиназой:
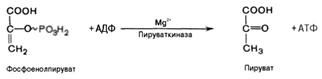
В результате одиннадцатой реакции происходит восстановление пировиноградной кислоты и образуется молочная кислота. Реакцияпротекает при участии фермента лактатдегидрогеназы и кофермента НАДН, образовавшегося в шестой реакции:

В пировиноградная кислота восстанавливается до молочной кислоты (лактата), поэтому в микробиологии анаэробный гликолиз называют молочнокислым брожением. Лактат далее ни во что не превращается, единственная возможность утилизовать лактат – это окислить его обратно в пируват.
Многие клетки организма способны к анаэробному окислению глюкозы. Для эритроцитов он является единственным источником энергии. Клетки скелетной мускулатуры за счет бескислородного расщепления глюкозы способны выполнять мощную, быструю, интенсивную работу, как, например, бег на короткие дистанции, напряжение в силовых видах спорта. Вне физических нагрузок бескислородное окисление глюкозы в клетках усиливается при – при различного рода анемиях, при нарушении кровообращения в тканях независимо от причины.
Гликолиз
Анаэробное превращение глюкозы локализуется в цитозоле и включает два этапа из 11 ферментативных реакций.
Первый этап гликолиза
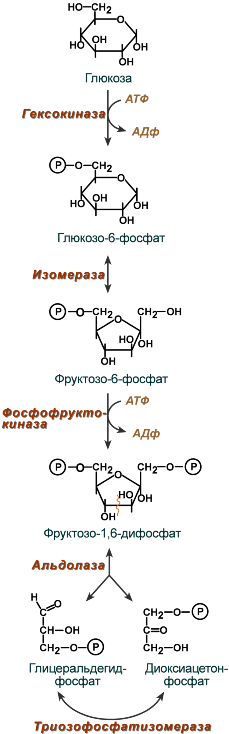
Первый этап гликолиза – подготовительный, здесь происходит затрата энергии АТФ, активация глюкозы и образование из нее триозофосфатов.
Первая реакция гликолиза сводится к превращению глюкозы в реакционно-способное соединение за счет фосфорилирования 6-го, не включенного в кольцо, атома углерода. Эта реакция является первой в любом превращении глюкозы, катализируется .
Вторая реакция необходима для выведения еще одного атома углерода из кольца для его последующего фосфорилирования (фермент ). В результате образуется фруктозо-6-фосфат.
Третья реакция – фермент фосфорилирует фруктозо-6-фосфат с образованием почти симметричной молекулы фруктозо-1,6-дифосфата. Эта реакция является главной в регуляции скорости гликолиза.
В четвертой реакции фруктозо-1,6-дифосфат разрезается пополам фруктозо-1,6-дифосфат- с образованием двух фосфорилированных триоз-изомеров – альдозы глицеральдегида (ГАФ) и кетозы диоксиацетона (ДАФ).
Пятая реакция подготовительного этапа – переход глицеральдегидфосфата и диоксиацетонфосфата друг в друга при участии . Равновесие реакции сдвинуто в пользу диоксиацетонфосфата, его доля составляет 97%, доля глицеральдегидфосфата – 3%. Эта реакция, при всей ее простоте, определяет дальнейшую судьбу глюкозы:
Второй этап гликолиза
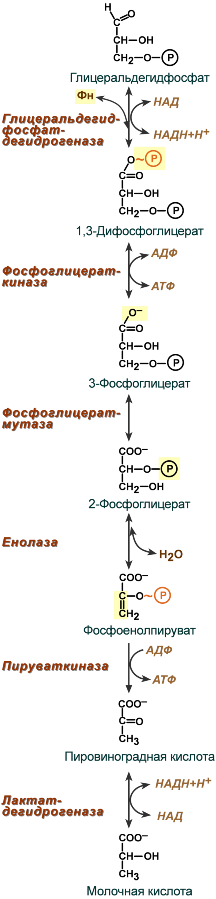
Второй этап гликолиза – это освобождение энергии, содержащейся в глицеральдегидфосфате, и запасание ее в форме АТФ.
Шестая реакция гликолиза (фермент ) – окисление глицеральдегидфосфата до кислоты и присоединение к ней фосфорной кислоты приводит к образованию макроэргического соединения 1,3-дифосфоглицериновой кислоты и НАДН.
В седьмой реакции (фермент ) энергия фосфоэфирной связи, заключенная в 1,3-дифосфоглицерате тратится на образование АТФ. Реакция получила дополнительное название – реакция субстратного фосфорилирования, что уточняет источник энергии для получения макроэргической связи в АТФ (от субстрата реакции) в отличие от окислительного фосфорилирования (использование энергии электрохимического градиента ионов водорода на мембране митохондрий).
Восьмая реакция – синтезированный в предыдущей реакции 3-фосфоглицерат под влиянием изомеризуется в 2-фосфоглицерат.
Девятая реакция – фермент отрывает молекулу воды от 2-фосфоглицериновой кислоты и приводит к образованию макроэргической фосфоэфирной связи в составе фосфоенолпирувата.
Десятая реакция гликолиза – еще одна реакция субстратного фосфорилирования – заключается в переносе макроэргического фосфата с фосфоенолпирувата на АДФ с образованием пировиноградной кислоты и АТФ.
Последняя реакция бескислородного окисления глюкозы, одиннадцатая – образование молочной кислоты из пирувата под действием . Важно то, что эта реакция осуществляется только в анаэробных условиях. Эта реакция необходима клетке, так как НАДН, образующийся в 6-й реакции, в отсутствие кислорода не может окисляться в митохондриях.
представляет собой высоко координированную и целенаправленную клеточную активность, обеспеченную участием многих взаимосвязанных ферментативных систем, и включает два неразрывных процесса и .
Он выполняет три специализированные функции:
Анаболизм
Анаболизм – это биосинтез белков, полисахаридов, липидов, нуклеиновых кислот и других макромолекул из малых молекул-предшественников. Поскольку он сопровождается усложнением структуры, то требует затрат энергии. Источником такой энергии является энергия .
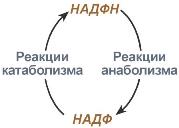
Цикл НАДФ-НАДФН
Также для биосинтеза некоторых веществ (жирные кислоты, холестерол) требуются богатые энергией атомы водорода – их источником является НАДФН. Молекулы НАДФН образуются в реакциях окисления глюкозо-6-фосфата в пентозосфатном пути или декарбоксилирования яблочной кислоты малик-ферментом. В реакциях анаболизма НАДФН передает свои атомы водорода на синтетические реакции и окисляется до НАДФ. Так формируется -цикл.
Катаболизм – расщепление и окисление сложных органических молекул до более простых конечных продуктов. Оно сопровождается высвобождением энергии, заключенной в сложной структуре веществ. Большая часть высвобожденной энергии рассеивается в виде тепла. Меньшая часть этой энергии “перехватывается” коферментами окислительных реакций и , некоторая часть сразу используется для синтеза .
Атомы водорода, высвобождаемые в реакциях окисления веществ, в основном используются клеткой по двум направлениям:
Необходимо заметить, что молекулы НАДФН могут идти не только на реакции анаболизма. Например, они активно привлекаются к реакциям антиоксидантной защиты для нейтрализации свободных радикалов, а в фагоцитирующих клетках, наоборот, требуются для синтеза супероксид анион-радикала, используются для нейтрализации аммиака в реакции синтеза глутамата в реакции восстановительного аминирования и в ряде других процессов.
Весь катаболизм условно подразделяется на три этапа, включающие реакции общих и специфических путей.
Происходит в кишечнике (переваривание пищи) или в лизосомах (самообновление клеток) при расщеплении уже ненужных или лишних молекул. При этом освобождается около 1% энергии, заключенной в молекуле. Она рассеивается в виде тепла.
Вещества, образованные при внутриклеточном гидролизе или проникающие в клетку из крови, на втором этапе обычно превращаются
Локализация второго этапа – цитозоль и митохондрии. На этом этапе выделяется около 30% энергии, заключенной в молекуле, и при этом запасается около 13% от всей энергии вещества (или примерно 43% от выделенной на этом этапе энергии).
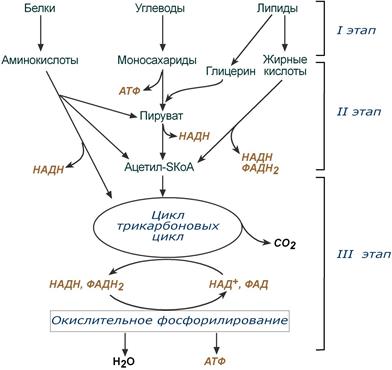
Схема общих и специфичных путей катаболизма(более подробная схема представлена здесь)
Под катаболизма понимают реакции, осуществляемые специфичными ферментами в специфичных, для разных классов веществ, реакциях 1 и 2 этапов. После того, как эти процессы закончатся, образуются пируват и ацетил-SКоА (в основном) и начинаются общие пути превращений. Подразумевается, что независимо от источника происхождения пирувата и ацетил-SKoA (из аминокислот, жирных кислот или моносахаридов) они попадают в катаболизма – 3 этап биологического окисления.
Третий этап
Все реакции этого этапа идут в митохондриях. Ацетил-SКоА (и кетокислоты) включается в реакции цикла трикарбоновых кислот, где углероды веществ окисляются до углекислого газа. Выделенные атомы водорода соединяются с НАД и ФАД, восстанавливают их и после этого НАДН и ФАДН2 переносят водород в цепь ферментов дыхательной цепи, расположенную на внутренней мембране митохондрий. Сюда же отдают свои атомы водорода молекулы НАДН и ФАДН2, образованные на втором этапе (гликолиз, окисление жирных кислот и аминокислот). В третьем этапе выделяется до 70% всей энергии вещества. Из этого количества усваивается почти две трети (66%), что составляет около 46% от общей. Таким образом, из 100% энергии окисляемой молекулы клетка запасает больше половины – 59%.
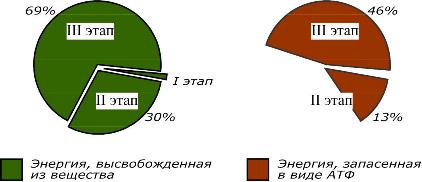
Соотношение выделенной и запасенной энергии при биологическом окислении
На внутренней мембране митохондрий в результате процесса под названием “” образуется вода и главный продукт биологического окисления – АТФ.
Роль АТФ
Энергия, высвобождаемая в реакциях катаболизма, запасается в виде связей, называемых макроэргическими. Основной и универсальной молекулой, которая запасает энергию и при необходимости отдает ее, является .
Все молекулы АТФ в клетке непрерывно участвуют в каких-либо реакциях, постоянно расщепляются до АДФ и вновь регенерируют.
Существует три основных способа использования АТФ:
Эти процессы вкупе с процессом образования АТФ получили название :
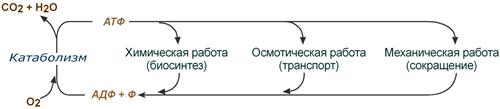
Кругооборот АТФ в жизни клетки
а) Анаэробные процессы высвобождения энергии. Анаэробный гликолиз. Иногда в случае отсутствия или недостаточного количества кислорода окислительное фосфорилирование становится невозможным. Но даже при таких условиях небольшое количество энергии может быть доставлено клеткам путем гликолитического расщепления углеводов, поскольку для химической реакции расщепления глюкозы до пировиноградной кислоты кислорода не требуется.
Это чрезвычайно неэкономичный путь метаболизма глюкозы, т.к. только 24000 калорий энергии, выделяемой при расщеплении каждой молекулы глюкозы, используется для образования АТФ, что составляет чуть больше 3% общего количества выделившейся энергии. Тем не менее, такой путь метаболизма, названный анаэробным энергообеспечением, в ситуации, когда кислород недоступен, доставляет энергию в течение нескольких минут, что может оказаться спасительным для клеток.
б) Образование молочной кислоты во время анаэробного гликолиза способствует выделению дополнительного количества энергии сверх анаэробного энергообеспечения. Согласно закону действующих масс, если количество образующихся конечных продуктов реакции приближается к средним значениям, обеспечиваемым реакцией, скорость реакций снижается практически до нуля. Конечными продуктами реакции гликолиза являются (для облегчения понимания просим вас изучить рисунок ниже):
(1) пировиноградная кислота;
(2) атомы водорода, присоединяющиеся к НАД+, образуя НАД-Н и Н+.
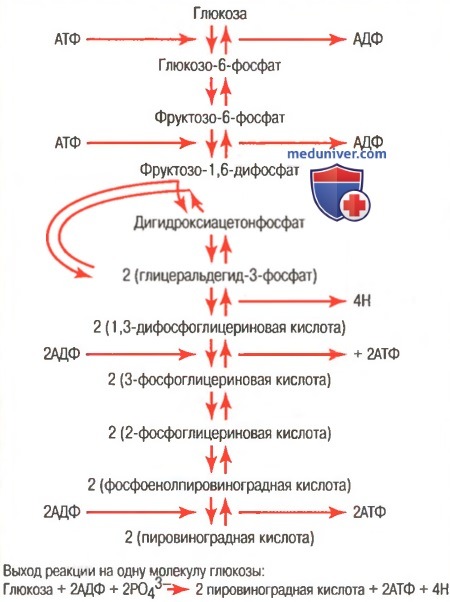
Последовательность химических реакций, ответственных за гликолиз
Образование обоих или одного из них останавливает процессы гликолиза и препятствует дальнейшему образованию АТФ. Если количество образовавшихся конечных продуктов реакции велико, они взаимодействуют друг с другом, образуя молочную кислоту в соответствии со следующей схемой реакции:
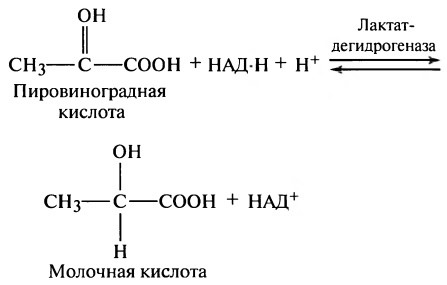
Таким образом, в анаэробных условиях большое количество пировиноградной кислоты превращается в молочную кислоту, которая легко диффундирует во внеклеточное пространство и даже внутрь некоторых менее активных клеток. Следовательно, молочная кислота представляет собой разновидность «водосточного колодца», в котором исчезают конечные продукты гликолиза, что позволяет гликолизу осуществляться дольше, чем это могло быть при отсутствии молочной кислоты.
Без этого превращения пировиноградной кислоты гликолиз мог бы осуществляться лишь в течение нескольких секунд. Вместо этого гликолиз продолжается в течение нескольких минут, снабжая организм значительным дополнительным количеством АТФ даже при отсутствии кислорода.
в) Обратное превращение молочной кислоты в пировиноградную, когда кислород становится доступным. Если кислород вновь становится доступным после периода анаэробного метаболизма, молочная кислота быстро превращается в пировиноградную кислоту, НАД-Н и Н+. Большие количества этих веществ немедленно окисляются, образуя значительное количество АТФ. Избыток АТФ может явиться причиной того, что более 75% пировиноградной кислоты вновь превращается в глюкозу.
Таким образом, большое количество молочной кислоты, которое образуется во время анаэробного гликолиза, не теряется организмом, т.к. если кислород вновь становится доступным, молочная кислота может подвергнуться обратному превращению в глюкозу или напрямую использоваться для получения энергии. Большая часть этих превращений осуществляется в печени, но в небольших количествах может происходить и в других тканях.
г) Использование сердцем молочной кислоты для получения энергии. Сердечная мышца обладает способностью превращать молочную кислоту в пировиноградную и затем использовать последнюю для получения энергии. В большинстве случаев это происходит при больших физических нагрузках, когда в кровь поступают значительное количество молочной кислоты из скелетных мышц и суммарно дает существенное количество энергии сердечной мышце.
Видео этапы, реакции гликолиза
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
– Также рекомендуем “Высвобождение энергии из глюкозы через пентозофосфатный цикл. Превращение глюкозы в жиры”
1. Образование АТФ через хемоосмотический механизм. Образование и синтез АТФ2. Синтез АТФ при расщеплении глюкозы. Выделение энергии из гликогена3. Анаэробный гликолиз. Молочная и пировиноградная кислота4. Высвобождение энергии из глюкозы через пентозофосфатный цикл. Превращение глюкозы в жиры5. Образование углеводов из белков и жиров. Регуляция глюконеогенеза6. Метаболизм жиров в организме. Транспорт липидов7. Свободные жирные кислоты. Транспорт свободных жирных кислот8. Липопротеины. Виды и физиология липопротеинов9. Обмен жиров и их депонирование. Жиры печени10. Расщепление жирных кислот. Окисление жирных кислот