алок чашечка короткая, растрескивается, рас слаиваясь на прикрепленные к плоду листья.
Разнообразие цветков у растений
Цветки растений могут быть однополыми или обоеполыми, а также могут располагаться на одной особи или на разных. Например, растения сочетания цветков на одной особи называют моноцезийными, а на разных особях — диоцезийными. Некоторые растения могут быть самоопыляемыми, если они обладают и мужскими, и женскими цветками на одной особи.
Многообразие сочетаний цветков
Многообразие сочетаний цветков на растениях поражает своим разнообразием: от кленов и ясеней с разноцветными цветками до гречихи с однополыми цветками. У многодомных растений сочетание цветков может быть даже более сложным, включая различные комбинации обоеполых, мужских и женских цветков на разных особях или даже на разных растениях.
Редкие случаи полной редукции цветков
Иногда наблюдаются случаи полной редукции половых функций цветка, приводящие к возникновению стерильных цветков. Эти цветки могут служить для привлечения опылителей, а обоеполые цветки обычно находятся в центре специализированных соцветий. Около 78% растений имеют обоеполые цветки, чего не скажешь о многодомных растениях, где сочетание цветков может быть еще более сложным.
Строение околоцветника
Околоцветник может быть простым, не разделенным на чашелистики и венчик, или же двойным, состоящим из чашелистиков и венчика. Чашелистики могут быть различной формы — от трубчатых до колокольчатых, и выполняют функцию защиты внутренних частей цветка. После цветения чашечка может остаться при плодах или же опадать, в зависимости от вида растения. Например, у фиалок чашечка короткая и растрачивается на прикрепленные к плоду листья.
Эта статья рассмотрела разнообразие цветков у растений, их сочетания и строение околоцветника. Важно помнить, что каждое растение имеет свои особенности в цветении, и изучение их помогает понять уникальные аспекты растительного мира.
Развитие цветкового покрова у зонтичных растений
Залиса (Physalis) трубочка разрастается, достигая длины 5—6 см и ширины до 5 см, становится пузыревидной и интенсивно-оранжевой по окраске. Иногда чашечка подобна венчику: крупная, ярко окрашенная. Отличаясь от венчика по форме и расположению (водосбор, фуксия и др.), функционально она может защищать венчик, на месте которого развиваются маленькие нектарники (купальница, морозник).
У многих зонтичных, сложноцветных, валериановых чашечка либо редуцирована, либо абортирована. Хохолок, способствующий разнесению плодов сложноцветных ветром, происходит от чашечки.
Особенности венчика цветка
Образует внутренний круг двойного околоцветника, отличаясь от чашечки более яркой окраской и более крупными размерами. Это самая заметная часть цветка, поэтому, говоря о размерах и окраске, имеют в виду обычно венчик. Лепестки могут быть свободные или срастаться на некотором протяжении, что приводит к образованию венчиков. В эволюционном плане, сростнолепестные венчики – более позднее приобретение по сравнению с раздельнолепестными.
Форма венчика и его симметрия
Длина трубки венчика может сильно различаться и обусловлена видом насекомого-опылителя. В зависимости от симметрии, венчик может быть:
- Актиноморфным (правильным) – если в нем можно провести несколько плоскостей симметрии.
- Зигоморфным (неправильным) – если в нем можно провести только одну ось симметрии.
- Асимметричным
Чашечка, венчик и цветок в целом обычно характеризуются явлением симметрии, причем симметрию цветка связывают с венчиком. Некоторые виды обладают дополнительными элементами, такими как чешуйки, зубчики и различные образования, привлекая насекомых для опыления. Лепестки цветков могут быть цельными, зубчатыми или рассеченными, создавая впечатление о многолепестности венчика.
Развитие цветкового покрова у зонтичных растений очень интересное и разнообразное явление, демонстрирующее удивительные адаптации к окружающей среде и процессам опыления.
Венчик цветка: его структура и функции
Если через венчик невозможно провести ни одной плоскости симметрии. Такие венчики характерны для очень немногих растений (канновые).
Зигоморфность и актиноморфность
Зигоморфность и асимметрия развились в процессе эволюции цветка в целях лучшего приспособления к опылителям-насекомым. Актиноморфные цветки более примитивны и встречаются у более древних семейств — лютиковых, розовых, лилейных.
Иногда симметрия венчика не совпадает с симметрией цветка (у пасленовых венчик актиноморфный, а весь цветок — зигоморфный). Переход от актиноморфности к зигоморфности проявляется уже в сем. лютиковых (аконит, живокость).
Окраска и бархатистость лепестков
Окраска лепестков венчика чаще всего определяется антоцианами, растворенными в клеточном соке. Желтая окраска обусловлена наличием антохлоров (георгины, мак) или хромопластов; белая — объясняется отражением всех световых лучей межклетниками, заполненными воздухом. Бархатистость лепестков зависит от мелких сосочков, имеющихся на эпидермальных клетках.
Функциональное назначение венчика
Функциональное назначение венчика заключается в защите андроцея и гинецея, в привлечении насекомых, способствующих перекрестному опылению.
Нектар и нектарники
Адаптация в процессе эволюции к определенным видам насекомых-опылителей обусловила образование шпорцев у цветков видов различных семейств. Шпорец возникает из лепестка или листочка простого околоцветника и представляет собой полый орган.
Нектарники — это экскреторные железки растений, выделяющие сахаристую жидкость — нектар. Форма нектарников весьма разнообразна, они могут быть различными по форме и местоположению в цветке. Нектарники встречаются на различных частях цветка, играя важную роль в привлечении опылителей.
В заключении, венчик цветка является важным элементом, способствующим опылению и размножению растений. Его структура и функции тщательно разработаны эволюцией для обеспечения успешного процесса опыления растений.
Разнообразие цветковых растений
Разнообразие нектарников у цветковых вели ко. Безусловно, их образование имело огромное значение в эволюции, главным образом в адапта ции к насекомоопылению. Образо вание нектара растениями имеет огромное значе ние для человека. На этой особенности растений основана целая отрасль хозяйственной деятельно сти — пчеловодство.
Андроцей
Совокупность тычинок в одном цветке назы вают андроцеем. Разнообразие андроцея чрезвычайно велико и его различают по многим признакам.
По числу тычинок
- Один тычинки — ятрышник
- Две тычинки — вероника
- Три ты чинки — касатик
- Четыре тычинки — губоцветные
- Пять тычинок — сложноцветные
- Шесть тычинок — лилейные
- Десять тычинок — мо тыльковые
- Много тычинок — лю тиковые
По срастанию
- Все ты чинки свободные — лютики
- Тычинки сросшиеся — камелия
По расположению
- Спирально — лютиковые
- Кругами — различные виды
Строение тычинки
Тычинка состоит из ниж ним концом, которой она прикреплена к цвето ложу, а на верхнем несет пыльник, содержащий пыльцу. Разделена на две половинки (теки), соединенные между собой связником, который является продолжением тычиночной нити. В ка ждой половинке пыльника расположены по два пыльцевых гнезда, в которых образуются микро споры.
Тычиночная нить
Тычиночная нить — это стерильная часть цветка. Она может быть прямой, изогнутой, из вилистой, скрученной, а ее поверхность голой, опушенной, волосистой. Длина колеблется в значительных пределах. Длинной тычиночной нитью считается та, которая длиннее пыльника, а короткой — та, которая короче пыльника.
В поперечном сечении тычиночная нить бывает округлой, плоской, широкой. Покры тая эпидермисом, она сложена паренхимными клетками, в центре проходит проводящий пучок.
Итак, разнообразие цветковых растений не только чрезвычайно интересно, но и имеет огромное значение в природе.
бывает самый различный по форме: длинный, если основание его длиннее пыльника (чистотел, рододендрон); с удлиненной верхуш кой, если она вытянута над пыльником (форзиция); с различными придатками.
Связник хорошо заметен, если пыльник рас сматривать со стороны, противоположной центру цветка.
Если пыльник обращен связником кнару жи цветка, говорят о его ориентации; если же к центру цветка — об ориен тации.
Многие ученые считают, что тычинки разви лись в результате редукции микроспорофиллов каких-то голосеменных предков. Следовательно, тычинки, обнаруживающие листовой характер, наиболее примитивны (у многоплодниковых). Примером могут служить тычинки дегенерии (Degeneria) из порядка магнолиецветных. Андроцей цветка этого растения состоит из 30—40 ши роких плоских тычинок, на нижней стороне ко торых попарно сближены микроспорангии. Дифференцировка на нить, пыльник и связник от сутствует. В процессе эволюции лентовидные ты чинки переходят в более специализированные, дифференцированные на нить, связник и пыль ник.
У барбарисовых до сих пор сохраняется часть связника над пыльником (надсвязник). Половин ки пыльника слились в один четырехгнездный пыльник, оказавшийся в результате редукции надпыльника на верхушке тычинки. Таким обра зом, верхушечное расположение надпыльника вторично.
Однако не все исследователи принимают та кую трактовку происхождения тычинок, полагая, что тычинки типа дегенерии вторичны, а первич ными Являются ветвистые тычинки (как у маль вовых, молочайных).
I I if’/
розы морщинистой; — фиалки; луи круглоголового; — клещевины; — шалфея; — вороньего глаза; — льна; — репы пыльник; — тычиночная нить; — связник; — нектарники)
МИКРОСПОРОГЕНЕЗ И МУЖСКОЙ ГАМЕТОФИТ
Наиболее существенной частью тычинок яв ляется пыльник, представляющий собой не сколько сросшихся микроспорангиев, располо женных на редуцированной до тычиночной нити листовой части микроспорофилла. Следователь но, тычинки — это микроспорофиллы. Пыльник
состоит из двух половинок—(греч. theke — хранилище, вместилище), соединенных связни ком. В каждой теке есть две полости (пыльцевые гнезда, мешки, камеры), в которых развивается пыльца. Обычно пыльник четырехтнездный, но иногда перегородка между гнездами разрушается и он становится двугнездным (орхидные, некото рые ластовневые и лавровые). Двугнездность мо жет развиться вследствие недоразвития второго гнезда в каждой половине. У некоторых растений пыльник может быть одногнездным (аризарум и некоторые эпакрисовые) в результате исчезнове ния и перегородок, и ткани связника. Иногда в каждой половинке может быть более двух гнезд, формирующихся в результате появления добавоч ных перегородок. Например, у бобовых в каждой половинке по четыре гнезда, а у омелы их насчи тывается около 50.
По морфологическому строению пыльники могут быть разнообразными. По типу прикрепле ния к тычиночной нити различают пыльники (связником пыльник прикреплен к цвето ложу); (пыльник с тычиночной ни тью соединен по всей длине пыльника); (если отклоняются от вертикальной оси); (соединяются с тычиночной нитью посередине подвижным сочленением) и т. д. Форма пыльников может быть линейной, ланце товидной, эллиптической, яйцевидной; поверх ность — гладкой (паслен), морщинистой, пузыр чатой (цикламен), опушенной (шлемник, барви нок, ломонос). Пыльники могут быть свободные, если не соприкасаются с соседними (лилия, тюльпан); сросшиеся боковыми сторонами (не дотрога) или приросшие к рыльцу (орхидные, некоторые колокольчиковые).
У некоторых растений не все тычинки спо собны развивать пыльцу в пыльниках, их назы вают или Они имеют форму нитей (аистник), бугорков и становятся лепестковидными (канны). У зимовника, купаль ницы они превратились в нектарники.
Развитие тычинок и пыльцы (микроспорогенез) происходит следующим образом (рис. 248).
Тычинки появляются на цветоложе в виде округлых бугорков. Их дальнейший рост начина ется с формирования пыльников, и позже путем интеркалярного роста вытягивается тычиночная нить, внутри которой к связнику проходит один проводящий пучок. Пучок окружен паренхимой, которая снаружи покрыта типичной эпидермой с устьицами. На самых ранних этапах формирова ния пыльника в его будущих четырех гнездах паренхимные клетки субэпидермального слоя в ре зультате периклинального деления формируют два слоя клеток археспория. Клетки внутреннего слоя в результате деления в радиальном и танген циальном направлениях формируют спорогенную ткань, из которой в дальнейшем образуются мик роспоры. Клетки наружного, слоя в результате пери- и антиклинальных делений образуют в свою очередь три слоя клеток, окру жающих клетки спорогенной ткани. Из них на ружный слой (субэпидермальный) в дальнейшем превратится в фиброзный слой, или Клетки его крупные, теряют содержимое, их обо лочки имеют сетчатое или спиральное утолще ние. Такая структура оболочки вызывает при подсыхании более сильное сокращение, чем у клеток эпидермы, что и обеспечивает вскрытие пыльника.
Клетки второго слоя, расположенные под эндотецием (их может быть несколько), обычно мелкие, разрушаются, и их содержимое использу ется на питание формирующейся пыльцы. Клет ки же самого внутреннего из трех слоев делятся, разрастаются, имеют густое содержимое. Они об разуют так называемый выстилающий слой, или тапетум. В процессе формирования пыльцы обо лочки клеток тапетума разрушаются, а их содер жимое вместе с содержимым граничащего с ним слоя клеток образует массу цитоплазмы с ядра-
Развитие пыльцевых гнезд
формирование; — однопыльцевое гнездо; эпидермис; субэпидермальный слой; • эндотеций; выстилающий слой; археспорий

ми — периплазмодий, расходуемый на питание пыльцы (рис. 249).
Клетки спорогенной ткани в результате деле ния образуют материнские клетки микроспор. Формирование микроспор из материнских клеток называют При этом диплоид ная материнская клетка делится мейозом. В ре зультате первого деления мейоза (редукционного) образуются две клетки уже с гаплоидным набо ром хромосом. Сразу же эти клетки делятся обычным митозом, из каждой образуются две клетки с гаплоидным набором хромосом. В итоге из диплоидной материнской клетки формируется
Строение вскрывшегося пыльника:
— пыльцевое гнездо; — связник; — проводящий пучок; — эпидермис; — эндотеций; — пыльцевые зерна — микроспоры
тетрада гаплоидных клеток. При этом деление может осуществляться по (клеточ ная оболочка возникает сразу же после первого деления, а затем после каждого второго деления в дочерних клетках) или по (перего родка после первого деления не образуется и ма теринская клетка расчленяется сразу же на четы ре части после обоих ядерных делений) типу. Стадия тетрады обычно кратковременна, и мик роспоры быстро обособляются друг от друга, а их цитоплазма формирует собственные оболочки. У вересковых, рогоза, росянки, элодеи микроспоры остаются в тетрадах, образуя сборные пыльцевые зерна; у мимозных пыльцевые зерна формируют более крупные скопления — до 64 клеток, а у не которых ластовневых, орхидных все микроспоры остаются в единой массе, называемой
Оболочка микроспор у немногих растений (ятрышник) остается тонкой, однородной. У мно гих же в результате утолщения изнутри и снаружи образуются две оболочки: внутренняя тонкая, со стоящая из пектиновых веществ, — и
наружная толстая, частично кутинизированная, обычно слабо опушенная — которая со держит нерастворимые в кислотах и щелочах уг леводы спорополленина. В свою очередь экзина имеет два слоя: наружный — (скулъптурированная часть) и внешний — (нескульптурированная часть). Строение внешнего слоя очень разнообразно и постоянно в пределах сис тематических групп, что широко используется в таксономии.
В экзине есть неутолщенные места или сквоз ные отверстия, через которые проникает при прорастании пыльцевая трубка. Эти места назы вают Расположение и форма апертур также разнообразны. Различают апертуры поляр ные, зональные (на экваторе), глобальные (рав номерно рассеянные по всей поверхности). По форме апертуры делятся на борозды и поры. Наиболее примитивными являются однобороздные пыльцевые зерна (голосеменные, магнолие вые). У большинства двудольных пыльца трехбороздная.
Размеры микроспор (пылинок) варьируют в за висимости от видовой принадлежности от 0,008 мм у фикуса (Ficus) до крупных, видимых простым глазом (до 0,2 мм) у тыквы. Форма пылинок ша ровидная, эллипсовидная, палочковидная, куби ческая, тетраэдрическая, нитевидная (рис. 250).
Морфологические признаки пыльцевых зерен широко используются в научных и практических целях (анализ меда, растительных остатков, в систематике) и послужили основанием для осо бого направления ботаники — палинологии. Пре красная сохранность пыльцы в ископаемых ос татках позволяет в результате анализа торфа уста новить систематический состав флоры предыду щих периодов. Такой метод исследования полу чил название спорово-пыльцевого анализа.
Содержимое микроспоры представляет собой густую протоплазматическую массу с большим запасом веществ — масел, Сахаров, крахмала и т. д. В микроспоре происходит развитие мужского гаметофита, которое сводится всего к одному де лению. Внутри микроспоры развивается система вакуолей и в пристенном слое происходит деле ние, в результате которого возникают две клетки:
или клетка пыльцевой трубки. Часто ее называют вегетативной, что позволяет гомологизировать ее с проталлиалышми клетками голосеменных, представляющих рудименты вегетативных клеток заростков. У покрытосеменных проталлиальные

Схема возникновения мужского гаметофита у сосны (верхний ряд) и у цвет кового растения (нижний ряд):
— проталлиалыше клетки; — спермагенная клетка; — клетка-ножка; — сифоногенная клетка; генеративные клетки
клетки утрачены и, следовательно, гаметофит достиг крайней редукции и состоит всего из двух клеток. Ядро генеративной клетки делится мито зом и из него образуются две мужские половые гаметы — Процесс формирования гамет (гаметогенез) происходит у многих растений еще в пыльнике, а у некоторых — после попадания пыльцы на рыльце пестика.
Сравнивая формирование гаметофита голо- и покрытосеменных, приходим к следующему вы воду: если у голосеменных он образуется в ре зультате четырех делений, то у покрытосеменных — в результате одного; у покрытосеменных отсутст вуют проталлиалыгые и антеридиальная клетки. Функцию спермагенной клетки выполняет гене ративная (рис. 251).
После созревания пыльцы в результате подсы хания фиброзного слоя происходит растрескива ние стенок пыльника и его вскрытие продольной трещиной, которая проходит вдоль перегородки между гнездами и вскрывает сразу оба гнезда. У манжетки, селезеночника вскрьшание пыльников происходит поперечной трещиной; дырочками на верхушке — у паслена, картофеля, грушанки; клапанами — у барбариса, лавровых.
При попадании в воду (за исключением рас тений, опыляемых водой) пыльца растений набу хает и лопается. Для предотвращения этого рас тения выработали многочисленные адаптации: поникшее положение цветка (ландыш, напер стянка, черника и др.); расположение цветков под листьями (липа, недотрога); закрывание цветков или соцветий (корзинок) во влажную погоду; расположение тычинок под защитой лепестков (мотыльковые, губоцветные, норичниковые) и т.д.
Гинецеем (греч. gyne— женщина и oikion — дом, жилище) называют совокупность плодоли стиков (мегаспорофиллов) в цветке, образующих одни или несколько пестиков. Это репродуктив ная часть цветка. Плодолистик, или мегаспорофилл, представляет собой листовую структуру, несущую семязачатки, и является составной ча стью пестика. На листовую природу плодолисти
ка обратил внимание еще И. Гёте, и в качестве доказательства этого обстоятельства обычно ука зывают на сходство плодолистика со свернув шимся листом (бобовые, некоторые лютиковые), наличие мезофилла, устьиц в эпидерме, сходство способа роста, отсутствие срастания краев плодо листика на самых начальных этапах развития у некоторых многоплодниковых и бобовых. Кроме того, при многочисленных тератологических слу чаях (уродствах) плодолистики превращаются в листовые структуры. В то же время ботаники не признают гипотезы возникновения плодолистика в результате метаморфоза вегетативных листьев цветковых растений. Наиболее признанным явля ется предположение, что плодолистики произош ли из мегаспорофиллов древних голосеменных. Начало вегетативным листьям и мегаспорофиллам, вероятно, дали недифференцированные лис товые органы папоротниковидных. Вероятнее всего, плодолистики возникли из открытых ме гаспорофиллов на той стадии развития, когда они были сложенными вдоль. Среди ньше живущих покрытосеменных есть виды с незамкнутыми плодолистиками (некоторые магнолиевые). На пример, у Degeneria плодолистик имеет ножку и сложенную вдоль пластинку, сросшуюся только в базальной части, а на большей части даже не со прикасающуюся краями. Семязачатки располо жены далеко от краев, а участки плодолистика между ними и краями сближаются, но не сраста ются. Функции рыльца выполняют поверхности по краям плодолистика, покрытые железистым опушением (рис. 252).
Пестик, возникающий из одного плодолисти ка (у бобовых), срастается по одному
Плодолистик Degeneria на двух стадиях развития

расположенному напротив средней жилки плодолистика. Если в цветке один или несколько отдельных пестиков, то гинецей называют (греч. арб — приставка, обозначающая отделение, и carpos — плод) и из каждого, пестика развивается плод (многие розоцветные, лютико вые). У большинства растений несколько плодо листиков срастаются и образуют один пестик, называемый (греч. kainos — новый) (рис. 253).
Гинецей, образованный тремя плодолистиками:
— апокарпный; — ценокарпный в разных степенях срастания плодоли стиков (3 — завязь; столбик; — рыльце)
Пестик расчленен на (нижняя разрос шаяся часть пестика, содержащая или мегаспорангии), (в апокарпном пестике) или (в ценокарпном, образую щемся при срастании стилодиев) и
Завязь от положения по отношению к другим частям цветка может быть верхней, нижней или полунижней (рис. 254). Верхняя завязь располага ется свободно на цветоложе, а все части цветка находятся под ней. Цветок в таком случае назы вается Нижняя завязь формирует ся в том случае, если все части цветка располага ются над завязью вследствие срастания их ниж них частей со стенками завязи (цветок называют При полунижней завязи части цветка срастаются со стенками завязи до полови ны последней, а завязь наполовину свободна.
Морфологическое разнообразие завязи вели ко. По форме завязь может быть шаровидной, яйцевидной, эллиптической, конусовидной, ци линдрической и т. д.; по внешнему виду — угло ватой или гранистой (от трехдо многогранной),
Положение завязи в цветке:
верхняя; 2, 3 — верхняя (или средняя); нижняя, образованная цвето ложем и плодолистиками; 5 — нижняя, образованная плодолистиками, срос шимися с нижними частями чашелистиков, лепестков и тычинок; — полу нижняя
крылатой, скрученной, с различными выростами. Кроме того, она может быть гладкой и морщини стой, голой и с различного рода опушением.
Внутри завязи в полости (камера, гнездо) за ключены семяпочки, следовательно, в отличие от голосеменных к ним непосредственно не могут попадать пыльцевые зерна. По числу гнезд завязь может быть одно-, двух-, трех-, четырех-, пяти- и многогнездной. По числу семязачатков завязь различают с одним, двумя, тремя и многими се мязачатками.
Место прикрепления семязачатка к завязи на зывают (лат. placenta — лепешка), а расположение плацент в завязи — В завязи различают два основных типа плацентации: семязачатки расположены на свободной внутренней поверхности (сусак, маг нолия), и — семязачатки расположе ны вдоль швов. Каждый из названных типов в свою очередь делится на отдельные формы.
Рыльце служит для улавливания пыльцы, и его морфологическое разнообразие связано с приспособлением к определенным типам опыле ния и конкретным опылителям. Оно может быть расположено на столбике и сидячее (если стол бика нет — у мака); по расположению на столби ке — верхушечное, боковое и поперечное; по размерам — большое (если его диаметр больше диаметра завязи — кувшинка белая, аспидистра), маленькое (кипрей), незаметное (лещина). Разли чают рыльце, в разной степени расчлененное на несколько равных или неравных частей, или ло пастей. Поверхность его может быть голой, с со сочками, с железками, опушенная, складчатая, бугорчатая, сухая и липкая.
Столбик, соединяющий завязь и рыльце, не является обязательной частью пестика, но у большинства растений присутствует. Верхняя его часть, располагающаяся под рыльцем, называется а нижняя, соединяющая его с завязью,-! В зависимости от расположения на завязи различают столбик или (если он расположен на апикальной части завязи); или (когда выходит сбоку завязи — земляника лесная), и (если расположен в центре завязи). По положению в пространстве столбик может быть прямостоячий, отклоненный, отопгутый, изогнутый, извилистый и т. д. Его толщина ко леблется в широких пределах. Часто в цветках растений одного вида встречается столбик разной длины, это явление называют или (у первоцвета есть длинно- и короткостолбчатые цветки). В ценокарпном ги нецее стилодий могут срастаться на разную дли ну, образуя часто один столбик. В таком случае по числу рылец можно определить число плодо листиков, из которых состоит гинецей (рис. 255).
Обсуждая происхождение плодолистика, мы отмечали как наиболее примитивный апокарп ный гинецей у Degeneria. Но неполное срастание плодолистиков встречается и среди представите лей других таксонов. Отчетливо выражены эпидермальные слои на поперечном срезе плодоли стика в месте соприкосновения краев у пионов,

Различные формы пестиков:
— завязь; — столбик; — рыльце
некоторых розоцветных, толстянковых. С утратой эпидермальньгх структур в процессе эволюции брюшной шов в месте срастания остается скорее воображаемой, чем реальной структурой. Наряду с редукцией брюшного шва наблюдается слияние брюшных пучков в общий пучок.
Рыльце в процессе эволюции постепенно ло кализуется в верхней части плодолистика, кото рая сама вытягивается и превращается в стилодий. Исходным, более примитивным считается плодолистик с нисходящим рыльцем и неоформ ленным стилодием. Развитие плодолистика с хо рошо развитым стилодием и локализованным в верхней его части рыльцем считается более про грессивным. Примитивные плодолистики харак терны для многих лютиковых. Основным преоб разованием внутренней части стилодия является специализация ткани в целях проведения пыль цевой трубки. Наиболее существенным преобра зованием гинецея в процессе эволюции стало возникновение ценокарпии и нижней завязи. Ценокарпный гинецей, как указывалось выше, возник в результате срастания плодолистиков. Срастание часто распространено только на завязь, при этом стилодии и рыльца остаются свободны ми (сложноцветные, гвоздичные). Количество рыльцев и стилодиев свидетельствует о количест ве плодолистиков. Во многих случаях срастание захватывает все части плодолистиков и образует ся столбик (норичниковые, бурачниковые, мно гие крестоцветные).
Прогрессивная роль ценокарпии доказывает ся прежде всего тем, что она имеет место в по давляющем большинстве сем. покрытосемен ных. Даже в наиболее примитивном сем. магно
лиевых есть виды со сросшимися плодолисти ками.
У нынеживущих растений различают три типа ценокарпного гинецея: синкарпный, паракарпный и лизикарпный.
сформировался в резуль тате бокового срастания апокарпных плодолисти ков (тюльпан, рябчик) и обычно имеет два гнезда
более. Семязачатки располагаются во внутрен них углах гнезд завязи, по краям завернувшихся внутрь плодолистиков, т. е. здесь типичная сутурального типа (яблоня, груша, лилейные).
образуется в результате срастания краями нескольких плодолистиков. В этом случае формируется одногнездная завязь. Образование ее могло произойти непосредствен но из еще не замкнутых плодолистиков или же в результате разъединения и выпрямления остав шихся сросшимися плодолистиков (мак, фиалка
др.). В паракарпном гинецее (париетальная — тыквенные). У многих растений в паракарпном гинецее плаценты силь но выпячиваются в полость завязи, формируя ложные перегородки, смыкающиеся или не смы кающиеся в центре (завязь мака состоит из 4—20 плодолистиков). Ложные перегородки могут воз никать и из выростов внутренней стенки завязи (у крестоцветных двугнездная завязь). Обычно в завязи паракарпного гинецея расположено много семязачатков, но у сложноцветных и злаков в ре зультате редукции их число уменьшается до од ной.
гинецей формируется в результа те срастания пестиков между собой боковыми

стенками плодолистиков, однако стенки затемзачаток гомологом мегаспорангия споровых рас
исчезают, за исключением сросшихся краев, не
сущих плаценты и называемых колонкой. Колони Семязачаток состоит из
ка на первый взгляд кажется продолжением оси(ядра), окруженного (по
цветка. Не исключена возможность и разрастаниякровами), которые на верхушке семязачатка не
цветоложа, но оно покрыто тканью*плодолистисмыкаются, образуя узкий канал —
ков. Такая плацентация называется (пыльцевход), через который пыльцевая трубка
или (гвоздика, первоцветные,проникает к зародьппевому мешку. Основание
гречишные). Иногда плацента сильно разрастаетнуцеллуса, от которого отходят интегументы, на
ся, заполняет гнезда завязи, и ее называют мяси
стой (пасленовые). В завязи может быть от одноплаценте с помощью (семяножки).
го до нескольких тысяч семязачатков (рис. 256).Место прикрепления семязачатка к семяножке
называют (рис. 257). Интегумент, по
крывающий семязачаток, может быть в единст
венном числе (роза, земляника, хмель), но покро
вов может быть и два (у большинства покрытосе
менных). В таком случае различают наружный,
или внешний, и внутренний интегумент. Иногда
интегументы отсутствуют, и тогда семязачаток
называется голым, без покровов (санталовые,
сих пор неясна. Наличие двух интегументов рас
сматривается как более примитивный признак.
иногда близкородственные таксоны раз
личаются по этому признаку. Так, у березовых один интегумент, а у граба, относящегося к этому семейству, — два; у осины один интегумент, а у многих видов этого рода — два.
Нуцеллус занимает центральную часть семяза-
Схемы эволюции основных типов гинецея:
плодолистики еще не замкнуты; апокарпный гинецей; синкарпный; паракарпный; 7, лизикарпный
Таким образом, исходным типом гинецея был апокарпный с большим количеством семяпочек в каждой одногнездной завязи, образованной од ним плодолистиком.
Одно из первых направлений эволюции — уменьшение числа плодолистиков до двух-трех и наконец до одного (бобовые, живокость, слива, вишня и др.) и числа семязачатков также до од ного (лютики, ветреницы). Второе направление эволюции — развитие ценокарпности, т. е. разви тие из апокарпного гинецея синкарпного, а из него — паракарпного и лизикарпного. В ценокарпном гинецее эволюционные преобразования шли по пути постепенной редукции числа плодо листиков (до двух) и числа семязачатков (до од ного).
МЕГАСПОРОГЕНЕЗ И ЖЕНСКИЙ ГАМЕТОФИТ
Семязачаток (семяпочка) — многоклеточное образование семенных растений, из которого раз вивается семя. Многие эмбриологи считают семя
Ортотропный семязачаток у горца (Polygonum
наружный интегумент,— внутренний интегумент, Я — нуцеллус;
зародышевый мешок; гипостаза; халаза; Ф— фуникулус;
С — синергиды;— антиподы; — вторичное адро;
чатка, и в нем развивается зародышевый мешок. В онтогенезе нуцеллус развивается первым и по морфологической природе является мегаспорангием, так как в нем расположен зародышевый мешок (мегаспора). Различают два типа нуцеллу са: тонкий, если он слабо развит, и тогда семяза чаток называют и толстый, если он хорошо развит, а семязачаток в этом слу чае называют
В процессе развития семязачатка у большин ства растений в его халазальной части обособля ются клетки, стенки которых сильно преломляют свет. Эту группу клеток называют роль которой окончательно не выяснена.
У многих покрытосеменных плацента образует вздутия в виде капюшона над нуцеллусом или сосочки из участков тканей семяножки, внутрен него интегумента или внутренней стенки завязи. Это так называемые которые растут по направлению к микропиле и обеспечивают проникновение пыльцевой трубки в зародыше вый мешок, ее рост и питание. У многих расте ний на семяпочке, а затем на семени развиваются мясистые образования — (присемянник, кровелька), частично или полностью прикры вающие семя и иногда ярко окрашенные (тисе, бересклет). В ткани ариллуса много Сахаров, мас ла, что способствует распространению семян жи вотными (копытень, чистотел и др.).
У покрытосеменных растений различают не сколько типов семязачатков. Еще К. Гебель (1932) в зависимости от положения нуцеллуса и микропиле по отношению к оси фуникулуса, от положения микропиле по отношению к базальной и апикальной частям завязи и степени изо гнутости нуцеллуса, а Боке (1959) в зависимости от положения проводящей системы и нуцеллуса по отношению к вертикальной оси семязачатка описали основные типы семязачатков (рис. 258).
Основные типы семязачатков:
орготропный; анатропный; J— гемитропный; — кампилотропный; амфитропный; зародышевый мешок
1. Ортотропный (атропный, прямой) — микро пиле и фуникулус расположены на противопо ложных концах оси семязачатка (крапивные, оре ховые, гречишные, ароидные).
2. Анатропный (обратный) — нуцеллус повер нут по отношению к прямой оси семязачатка на 180 °, вследствие чего микропиле и фуникулус расположены рядом (у большинства покрытосе менных).
3. Гемитропный (полуповернутый) — семязача ток повернут на 90 °, вследствие чего микропиле и нуцеллус расположены по отношению к фуникулусу под углом 90 ° (некоторые норичниковые, первоцветные).
нуцеллус изогнут толь ко микрополярным концом к основанию семя зачатка, поэтому микропиле расположено рядом с фуникулусом (мальвовые, бобовые, резедо вые).
5. Амфитропный (двусторонне изогнутый) — ну целлус изогнут в виде подковы и микропиле вследствие этого расположено рядом с фуникулу сом (тутовые, ладанниковые и др.).
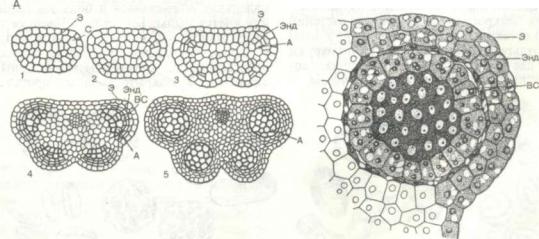
Развитие семязачатка и мегаспорогенез. Семя зачаток возникает на плаценте в форме неболь шого бугорка, сложенного однородными меристематическими клетками. В результате антикли нального деления клеток эпидермы нуцеллуса и периклинального деления клеток субэпидермального и более глубоколежащих слоев осуществля ется рост семязачатка. После достижения семяза чатком определенной величины в его основании возникают в виде кольцевых валиков интегументы, а в апексе семязачатка вычленяются одна или несколько клеток археспория (эти клетки разви ваются из субэпидермальной клетки нуцеллуса под пыльцевходом). Археспорий может быть мно гоклеточным, что является примитивным призна ком, но более распространен одно-, двухклеточный археспорий. Эти клетки более крупные и с густой цитоплазмой.
В крассинуцеллятных семязачатках археспориальная клетка (или одна из них) делится на пер вичную (кроющую) клетку и которая и является материнской клет кой мегаспоры. В тенуинуцеллятных семязачатках археспориальная клетка становится предшествен ницей мегаспоры.
Формирование мегаспоры из архегониальной или спорогенной клетки в результате мейоза на зывают
Археспориальная, или спорогенная, клетка делится путем мейоза. Последовательно происхо дят два деления: первое редукционное, а затем два гаплоидных ядра делятся типичным митозом. В итоге формируются четыре гаплоидные клетки, расположенные, как правило, по оси семязачатка. Обычно нижняя (реже верхняя или промежуточ ная) клетка разрастается, а три остальные дегене рируют. Эта разросшаяся клетка и является ме гаспорой, которая дает начало зародышевому меш ку, называемому Такой путь характерен для большинства покрытосеменных (рис. 259).
Однако у растений наблюдаются многочис ленные отклонения от типичного пути формиро вания зародышевого мешка.
1. У лилейных, рясок, кермеков археспориаль ная клетка становится зародышевым мешком без предварительного мейоза, т. е. он изначально ди плоидный. Редукция числа хромосом происходит уже при делении в прорастающем зародышевом мешке.
2. При делении археспориальной клетки фор мирование клеточных перегородок может быть подавлено и образуется клетка с четырьмя сво бодными ядрами. В этом случае формируется четырехспоровый зародышевый мешок (тюльпан, лилия, майник).